Спорная книга: Станислав Дробышевский, «Достающее звено»
Станислав Дробышевский. Достающее звено. Книга первая. Обезьяна и все-все-все
М.: АСТ. Corpus, 2017
Станислав Дробышевский. Достающее звено. Книга вторая. Люди
М.: АСТ. Corpus, 2017
Елена Макеенко в обзоре «Что читать этим летом: 10 книг в разных жанрах» («Esquire») кратко рассказывает, «о чем кино» — впрочем, счастливо избегая спойлеров: «Внушительный двухтомник, отвечающий на все вопросы антропогенеза, — звучит страшновато, но на деле это отличный способ провести свое читательское лето с пользой и удовольствием. Все потому, что его автор Станислав Дробышевский — ведущий российский антрополог, преподаватель МГУ, научный редактор сайта Антропогенез.ру и в хорошем смысле поп-звезда от науки. Если кто-то и мог соблюсти научную ценность и увлекательность повествования об эволюции на одинаково высоком уровне, то вот он, знакомьтесь. В двух томах — “Обезьяны и все-все-все” и “Люди” — Дробышевский подробно и в меру иронично излагает содержание предыдущих серий происхождения человека, рассказывает обо всех возможных предках Homo Sapiens, объясняет, почему и как человек родился именно в семействе гоменидов, и даже предсказывает наше будущее как вида с точки зрения антропогенеза».
Особо подчеркивает легкость пера Дробошевского Галина Юзефович в обзоре «Семь отличных книг в жанре нон-фикшн, чтобы с пользой провести лето» («Медуза»): «Двухтомник антрополога Станислава Дробышевского — книга, из которой можно долго (возможно, бесконечно) черпать смешные байки и исторические параллели и которую хочется, едва дочитав, купить в подарок другу — разумеется, не бескорыстно, а чтобы потом обсудить. Написанная с ироничной легкостью, она заставляет забыть о том, что ее автор — один из ведущих специалистов в области антропогенеза, крупнейший исследователь человека как биологического вида.
“Достающее звено” — это одновременно и книга о том, как один вид древних гоминид эволюционировал в самое высокоразвитое существо на земле, и планетарного масштаба семейная сага, и обстоятельный рассказ о том, как человек познавал собственную родословную. Если вам интересно, у кого больше шерсти — у нас или у шимпанзе, как размер зубов связан с интеллектом и почему идея происхождения человека от обезьяны до сих пор сталкивается с сильнейшим сопротивлением, книга Дробышевского будет для вас идеальным летне-познавательным чтением. Впрочем, если все это вас не интересует, все равно попробуйте почитать “Достающее звено” — скорее всего, Дробышевский сумеет вас переубедить».
Сергей Кумыш в обзоре «Два Чарльза и достающее звено: пять самых интересных книжных новинок июня» («Posta Magazine») акцентирует внимание на том, что двухтомник Дробышевского адресован далеко не только хардкорным читателям научпопа: «“Об антропогенезе человеческим языком” — в принципе, на этом максимально кратком пересказе двухтомника Станислава Дробышевского “Достающее звено” можно было бы остановиться. Люди, интересующиеся вопросом, наверняка книгу прочтут. Люди, смутно представляющие себе, что такое антропогенез, скорее всего, не дочитают до конца даже этот абзац. Но если вы все еще с нами, то вот, из первой главы: “В приложении к человеку теория эволюции называется антропогенезом”. На самом деле автору “Достающего звена” удается не просто максимально быстро ввести неподготовленного читателя в курс дела понятными, в лучшем смысле доступными словами, но, что гораздо важнее (и, учитывая специфику текста, куда сложнее), на протяжении всей книги удерживать читательское внимание на одном и том же уровне, и в итоге подарить ни с чем не сравнимое счастье владения фактом людям, от академического мира безнадежно далеким. Этот обстоятельный рассказ о происхождении человека, по объему не уступающий “Властелину колец”, на поверку оказывается чтением не менее захватывающим».
О хоббитах вспоминают и Полина Бояркина, Анастасия Сопикова и Анастасия Оленева в обзоре «Книга в дорогу. Часть 1» («ПРОчтение»): «Теория эволюции человека называется антропогенезом. Именно ей и посвящена книга биолога и популяризатора науки Станислава Дробышевского. По названиям томов — “Обезьяны и все-все-все” и “Люди” сразу становится понятно, кто в них главные герои. Прелесть научпопа в том, что он написан для неопытных читателей. Автору этой книги очень быстро удается ввести неподготовленных в курс дела и во многом благодаря легкому и метафорическому языку постоянно поддерживать интерес к чтению. Особенно заумные места можно пропускать, тогда даже останется время, чтобы поспать. Или, если будет не оторваться, дочитывать во время пересадок. Сергей Кумыш в статье на Posta Magazine по объему и увлекательности сравнил “Достающее звено” с “Властелином колец”, отведя в книге Дробышевского не последнее место хоббитам, — весьма высокая оценка художественных достоинств книги».
И наконец Владислав Толстов в материале «Книги о науке: хочу все знать» (
Ранее в рубрике «Спорная книга»:
• Джулиан Феллоуз, «Белгравия»
• Мария Галина, «Не оглядываясь»
• Амос Оз, «Иуда»
• А. С. Байетт, «Чудеса и фантазии»
• Дмитрий Глуховский, «Текст»
• Майкл Шейбон, «Лунный свет»
• Сборник «В Питере жить», составители Наталия Соколовская и Елена Шубина
• Владимир Медведев, «Заххок»
• Ю Несбе, «Жажда»
• Анна Козлова, «F20»
• Хелен Макдональд, «Я» — значит «ястреб»
• Герман Садулаев, «Иван Ауслендер: роман на пальмовых листьях»
• Галина Юзефович. «Удивительные приключения рыбы-лоцмана»
• Лев Данилкин. «Ленин: Пантократор солнечных пылинок»
• Юрий Коваль, «Три повести о Васе Куролесове»
• Андрей Рубанов, «Патриот»
• Шамиль Идиатуллин, «Город Брежнев»
• Фигль-Мигль, «Эта страна»
• Алексей Иванов, «Тобол. Много званых»
• Владимир Сорокин, «Манарага»
• Елена Чижова, «Китаист»
«Достающее звено.
 Книга первая. Обезьяны и все-все-все» Дробышевский Станислав Владимирович — описание книги | Элементы (Россия)
Книга первая. Обезьяны и все-все-все» Дробышевский Станислав Владимирович — описание книги | Элементы (Россия) Путь человека
Кто такие шерстокрылы, кому легче выжить, травоядным или хищникам, как уровень развития социума зависит от размера мозга, какими были первые доисторические карты, почему неандертальцы храпели, а кроманьонцы спали тихо, как растения научились убивать свои клетки?
Замечательный лектор, преподаватель кафедры антропологии биологического факультета МГУ, соучредитель и бессменный научный редактор легендарного портала Антропогенез.ру, неутомимый популяризатор науки Станислав Владимирович Дробышевский в своей новой книге расскажет все о том, как зарождалась, развивалась и видоизменялась жизнь на Земле.
«Обезьяны и все-все-все» — первая из двух книг фундаментальной работы «Достающее звено», в которых Дробышевский рассказывает, как появлялся и утверждался в этом мире человек, и почему то самое мистическое «недостающее звено» на самом деле уже давно было найдено.
В книге читатель найдет множество разных рубрик, например, «Уголки занудства», в которых во всех терминологических тонкостях и со всей научной серьезностью описаны те или иные важные факты и открытия, так или иначе связанные с историей антропогенеза. Раздел «Минутка фантазии», напротив, приглашает расслабиться и помечтать о том, какую цивилизацию могли бы создать гигантопитеки, или как человек мог бы в процессе эволюции получить 14 конечностей. Рубрика «Кстати, о…» раскроет некоторые удивительные факты, например, о том, почему пересадки печени, сердца и костного мозга от павиана человеку проходили успешно, а вот пересадка почек влекла за собой смерть пациента через несколько месяцев.
Великолепный юмор, отточенный стиль, легкость изложения даже самого сложного научного материала, огромное количество интересных фактов и достоверных историй – эта книга ответит на все ваши вопросы об эволюции и развитии жизни на земле.
Достающее звено. Кн.2. Люди (Дробышевский, С.)
Дробышевский, С.Станислав Дробышевский в своей новой книге расскажет все о том, как зарождалась, развивалась и видоизменялась жизнь на Земле. «Обезьяны и все-все-все» — первая из двух книг фундаментальной работы «Достающее звено», в которых Дробышевский рассказывает, как появлялся и утверждался в этом мире человек, и почему то самое мистическое «недостающее звено» на самом деле уже давно было найдено.
Полная информация о книге
- Вид товара:Книги
- Рубрика:Антропология
- Целевое назначение:Научно-популярное издание для взрослых
- ISBN:978-5-17-113935-3, 978-5-17-114004-5
- Серия:Несерийное издание
- Издательство: АСТ, CORPUS
- Год издания:2019
- Количество страниц:588
- Тираж:2000
- Формат:60х90/16
- УДК:572
- Штрихкод:9785171139353
- Переплет:в пер.

- Сведения об ответственности:Станислав Дробышевский
- Вес, г.:643
- Код товара:14537
«Достающее звено». Книга первая
Двухтомное сочинение Станислава Дробышевского «Достающее звено» вошло в шорт-лист премии научно-популярной литературы «Просветитель» 2017 года. В нем рассказывается об эволюции всего живого — первый том называется «Обезьяны и все-все-все», а второй — «Люди». Издание N + 1 предлагает своим читателям познакомиться с фрагментом из первого тома книги Дробышевского. И не забывайте, что ближе к финалу премии (16 ноября) мы запустим в паблике ВК «Образовач» голосование, в ходе которого вы сможете проголосовать за наиболее понравившуюся вам книгу из шорт-листа.
Звено 1: появление клеточного ядра
(протерозой, 1,9±2,4 млрд лет назад)
Первые ядерные организмы — эукариоты — возникли 1,9–2 млрд лет назад. Одной из наиболее аргументированных гипотез является симбиотическая. Собственно, симбиоз оказался итогом несварения: одна клетка глотала другую, но не растворяла, а использовала в мирных целях. Видимо, ядро было когда‑то археей, а из бактерий получились митохондрии.
Одной из наиболее аргументированных гипотез является симбиотическая. Собственно, симбиоз оказался итогом несварения: одна клетка глотала другую, но не растворяла, а использовала в мирных целях. Видимо, ядро было когда‑то археей, а из бактерий получились митохондрии.
Ядро — очень важный элемент клетки. Оно защищает ДНК, что повышает надежность сохранения информации и дает возможность разнообразить биохимические процессы. В цитоплазме бактерий ДНК неуютно, злые ферменты того и гляди норовят порвать ее в клочья. Кроме прочего, ДНК — это кислота, так что реакции с образованием щелочей бактериям противопоказаны, иначе ДНК нейтрализуется и выпадет в виде соли. Да и вообще, много ли проведешь противоречащих друг другу реакций в одной тесной бактериальной клетушке? Поэтому и ДНК у бактерий обычно маленькая, много генов ей и не надо. И форма у нее кольцевая — так ферментам труднее ухватиться, нет кончика, с которого бы можно было начать есть бедную молекулу. Но опять же кольцо не сделаешь слишком длинным — вырастет вероятность разрыва.
Другое дело в ядре! Любо там, тишь, благодать, примиренье. ДНК надежно спрятана за ядерной мембраной. Теперь ее можно раскольцевать в длинные хромосомы, а их самих напихать в клетку сколько душе угодно (у папоротника‑ужовника их 1260 в каждой клетке). Беспрецедентное увеличение генетического материала позволяет ввести в эксплуатацию новые, невиданные доселе обменные процессы, усложнить их, что в будущем станет залогом сначала колониальности, а после и многоклеточности.
Все опасные обменные процессы идут снаружи, в цитоплазме, да еще разнесены по отдельным пузырькам. С этого момента в полные права вступает “пузырькование”; через это странное слово можно определить всю сущность жизни. Жизнь как пузырькование пузырьков в пузырьках, емкостей, без конца вдавливающихся друг в друга и выпучивающихся наружу: на уровне клетки, эмбриональных бластул и гаструл, мозговых пузырей и зачатков органов чувств, трофобластов, амнионов и желточных пузырьков, мозговой, грудной и брюшной полостей, полых внутренних органов и даже вне организма — нор, гнезд и домов. Но стоп! Мы забежали слишком далеко вперед. В протерозое дело ограничилось лишь ядром и митохондриями.
Но стоп! Мы забежали слишком далеко вперед. В протерозое дело ограничилось лишь ядром и митохондриями.
ДНК эукариот защищена ядерной мембраной от вторжения извне — вирусов и горизонтального переноса генов от других организмов, что можно рассматривать как прогресс, так как позволяет сохранить свое “я”. Но изменчивость бактерий — в своем роде залог их успеха, благодаря ей они могут очень быстро приспосабливаться к новым условиям. Получив стабильность, мы — эукариоты — утеряли пластичность.
Митохондрии синтезируют АТФ, что дает огромные энергетические возможности. Энергию можно запасать по‑разному: в жирах, углеводах, белках, но АТФ — лучшая батарейка, хотя и недолговечная. Первые эукариоты, поглотившие бактерий, умевших синтезировать АТФ, получили такое преимущество, что все другие тут же проиграли эволюционную гонку. Сейчас на планете нет организмов, живущих без АТФ. Правда, остается под вопросом, существовали ли предки эукариот, не умевшие синтезировать АТФ, или они утеряли эту способность, положившись на новообретенные генераторы. В любом случае из поглощенных бактерий получились митохондрии, у них и до сих пор имеется бактериальная кольцевая ДНК и собственные рибосомы, они делятся самостоятельно и живут как внутриклеточные симбионты, причем в одной нашей клетке их может быть до двух тысяч. Но часть своих генов они сдали на хранение в более надежное ядро, так что теперь не так легко понять, где граница между собственно нами и нашими жильцами. Обретение митохондрий преобразило жизнь эукариот, благодаря им мы можем активно двигаться. Например, ими насыщена красная скелетная мускулатура, скажем прямые мышцы спины, держащие нас вертикально. Иначе говоря, метаболические нужды протерозойских одноклеточных обеспечили наше прямохождение.
Звено 2: аэробная атмосфера — многоклеточность и твердые части тела
(поздний протерозой и ранний кембрий, 850–540 млн лет назад)
Докембрийский мрак скрывает много тайн. Как возникли первые нервные клетки? Как появилось разделение на два пола? Слишком много гипотез, слишком мало фактов. Но все же о некоторых вещах мы можем говорить достаточно уверенно.
Но все же о некоторых вещах мы можем говорить достаточно уверенно.
В катархее, архее и начале протерозоя в атмосфере было много углекислого газа, метана, сероводорода, аммиака и водяных паров, но почти не было кислорода. Примерно 2,5 млрд лет назад или даже раньше цианобактерии начали производить свободный кислород, но он поглощался горными породами и шел на образование озонового слоя. Кстати, спасибо им за это, без него нам было бы нелегко, ведь озоновый слой защищает нас от космической радиации. Ко времени 850–600 млн лет назад упорные цианобактерии нафотосинтезировали катастрофически много кислорода и отравили им всю атмосферу (справедливости ради надо сказать, есть версия, что кислород сам высвободился из мантии Земли, без помощи бактерий). Для преобладавших в тогдашних экосистемах анаэробов (организмов, живущих в бескислородной среде) кислород был страшным ядом. По всей видимости, это привело к глобальному вымиранию. Приятно сознавать, что не только человек способен так загадить окружающую среду, что потом сам не может в ней жить. Мы же — потомки тех редкостных протерозойских аэробов, которые поначалу с трудом привыкали к ужасному яду в недрах цианобактериальных матов, зато потом попали просто в райские условия.
Кислородный обмен — хорошая вещь. С его помощью можно получать гораздо больше энергии, чем при анаэробном существовании. А с помощью этой энергии можно делать новые чудесные вещи, например увеличивать размеры тела и синтезировать новые вещества. В большей клетке можно хранить большее количество ДНК, а за счет этого усложнить свое строение.
В числе прочего появляется возможность стать многоклеточным. Жить толпой надежней и веселей, неспроста еще на уровне бактерий совершались попытки в этом направлении. Но истинно многоклеточное существо внутренне противоречиво. С одной стороны, все его клетки содержат одинаковую генетическую информацию, с другой — работать в разных клетках она должна по‑разному. Основная проблема возникает с размножением. С появлением многоклеточности в мире появились дряхлость и смерть. Одноклеточные не умирают от старости — они могут погибнуть лишь от случайности, голода или хищника. Главная цель любой приличной клетки — стать двумя клетками, жить вечно. Даже если такие организмы соберутся в кучку и станут как‑то общаться между собой, они еще не станут многоклеточным организмом, их можно назвать лишь колонией — бесформенной кучей одиночек‑эгоистов. В истинно многоклеточном общежитии нужен строгий контроль за размножением, большинство клеток гарантированно умрут, а потенциальным бессмертием обладают лишь гаметы — половые клетки. Убедить рядовую — соматическую — клетку не размножаться, а жить и добровольно сгинуть для обеспечения бессмертия гаметы очень трудно, для этого нужно много сложных генов (притом что гены во всех клетках одни и те же), а их можно хранить лишь в большой клетке с ядром. Поэтому бактерии так никогда и не смогли стать многоклеточными, а некоторые эукариоты таки преодолели сложность.
С появлением многоклеточности в мире появились дряхлость и смерть. Одноклеточные не умирают от старости — они могут погибнуть лишь от случайности, голода или хищника. Главная цель любой приличной клетки — стать двумя клетками, жить вечно. Даже если такие организмы соберутся в кучку и станут как‑то общаться между собой, они еще не станут многоклеточным организмом, их можно назвать лишь колонией — бесформенной кучей одиночек‑эгоистов. В истинно многоклеточном общежитии нужен строгий контроль за размножением, большинство клеток гарантированно умрут, а потенциальным бессмертием обладают лишь гаметы — половые клетки. Убедить рядовую — соматическую — клетку не размножаться, а жить и добровольно сгинуть для обеспечения бессмертия гаметы очень трудно, для этого нужно много сложных генов (притом что гены во всех клетках одни и те же), а их можно хранить лишь в большой клетке с ядром. Поэтому бактерии так никогда и не смогли стать многоклеточными, а некоторые эукариоты таки преодолели сложность.
Правда, и у многоклеточных бывают рецидивы, когда клетка “вспоминает”, что она свободна, никому ничего не должна и может делиться сколько влезет — тогда возникает рак. Причина — мутация, поломка генов‑ограничителей, держащих эгоизм в узде.
Получив многоклеточность, мы стали большими и сложными, но потеряли индивидуальное бессмертие, да еще получили в нагрузку шанс умереть от собственных клеток, стремящихся к бессмертию. Адекватна ли цена?..
Многоклеточность бывает разная. Первые опытные образцы, судя по современному трихоплаксу Trichoplax adhaerens, больше напоминали кляксу. Потом тело стало шариком; затем оно завернулось кувшинчиком, причем двухслойным — это уровень кишечнополостных и плоских червей. Но такая форма имеет маленький недостаток: вход и выход из пищеварительной системы — это одно и то же отверстие. Стало быть, нельзя есть непрерывно, а ведь хочется! Более того, такой пищеварительный тракт невозможно дифференцировать на части. Посему величайшим достижением неких протерозойских животных стало обретение анального отверстия.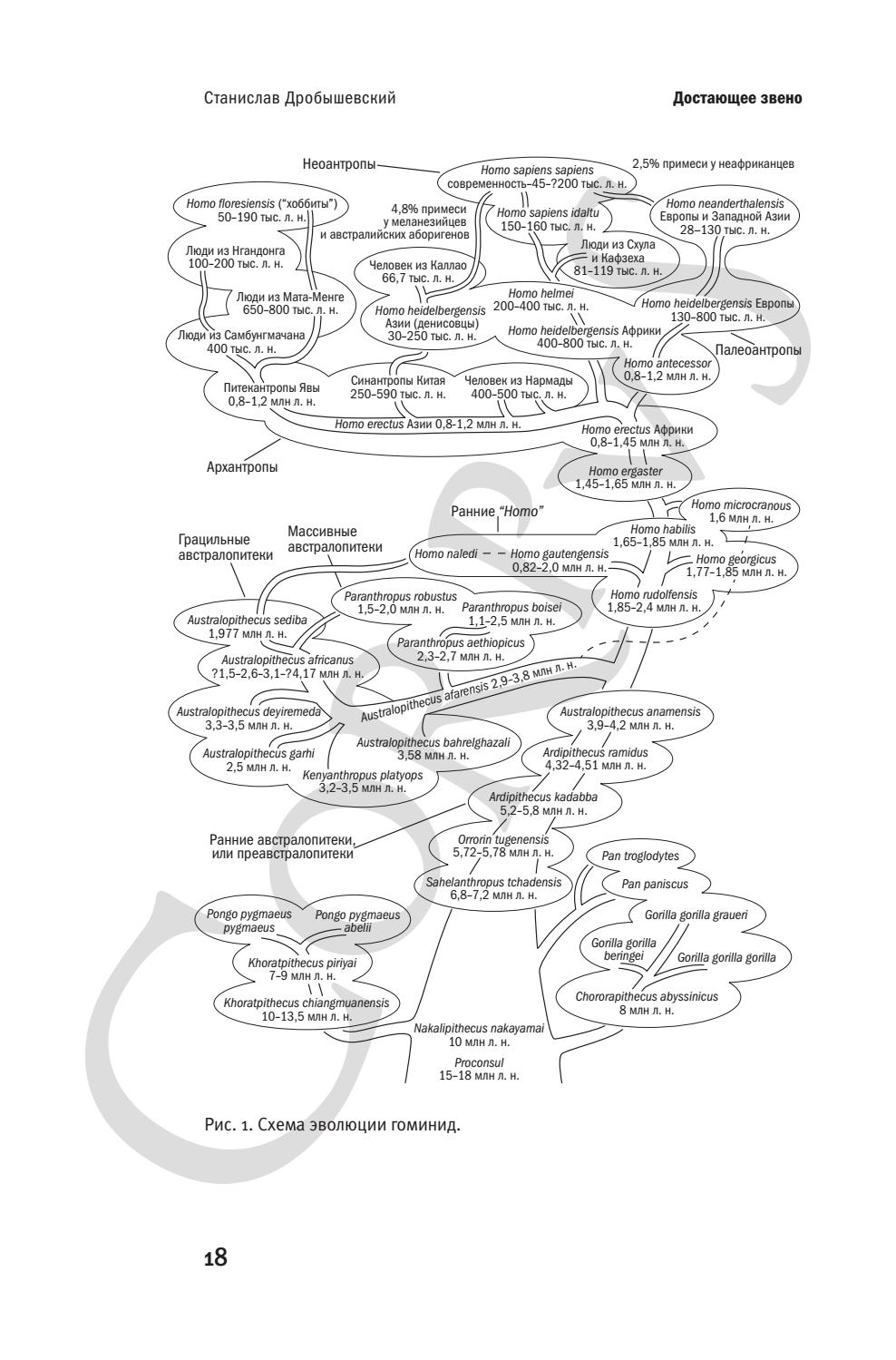 Тело преобразовалось из кувшина в трубку (каковой и пребывает доселе), теперь стало возможно лопать сколько влезет, а пищеварительный тракт — подразделить на сегменты: глотку, пищевод, желудок, переднюю кишку, заднюю кишку. Конечно, не все эти отсеки возникли одновременно, но главное было начать. А в разных отделах можно выделять разные ферменты, переваривать разные вещества, лучше их усваивать, а через то — повышать обмен веществ.
Тело преобразовалось из кувшина в трубку (каковой и пребывает доселе), теперь стало возможно лопать сколько влезет, а пищеварительный тракт — подразделить на сегменты: глотку, пищевод, желудок, переднюю кишку, заднюю кишку. Конечно, не все эти отсеки возникли одновременно, но главное было начать. А в разных отделах можно выделять разные ферменты, переваривать разные вещества, лучше их усваивать, а через то — повышать обмен веществ.
История возникновения пищеварительной системы каждый раз повторяется в нашем эмбриогенезе: сначала из бластулы впячиванием стенки внутрь образуется гаструла с одним отверстием — бластопором — наружу и полостью — гастроцелью — внутри, будущей пищеварительной системой. Фактически это уровень гидры или медузы. Потом с противоположной бластопору стороны вдавливается второе отверстие и соединяется с гастроцелью — это уровень круглых червей (условно, конечно). Человек — тоже трубка. Из сего, кстати, следует интересный казус: полость пищеварительной системы — это на самом деле внешняя среда по отношению к человеку, а бактерии‑симбионты, сидящие в нашем кишечнике, сидят в реальности снаружи от нас. Они и рады бы стать паразитами, да иммунная система внутрь не пускает. Потому же так просто решается загадка, столь мучащая многих школьников, постигающих анатомию: почему многие железы внешней секреции открывают свои протоки в желудок и кишечник? Вроде же они внутри человека? На самом деле протоки открываются вполне себе наружу, просто “наружа” завернута в нас и даже проходит сквозь нас.
Кстати, о ртах…
Многоклеточные животные в первом приближении делятся на две масштабные группировки — первично- и вторичноротые. К первым относятся, например, моллюски и членистоногие, ко вторым — иглокожие и хордовые. У первичноротых отверстие, появляющееся у гаструлы, становится в последующем ртом, а возникающее позже — анальным. У вторичноротых все наоборот, первым закладывается именно анальное отверстие. Долгие годы зоологи и эмбриологи спорили: свидетельствует ли такая разница о независимом происхождении этих групп, или она второстепенна. Точку в дискуссии поставило открытие Hox-генов — особого семейства генов, определяющих закладку переда, середины и зада. Оказалось, что эти гены мало того что весьма схожи от червей до человека, так они еще и жутко консервативны. Различия же первично- и вторичноротых определяются только разницей в последовательности формирования в общем-то одинаковых частей. Мы начинаем формироваться сзади наперед, а мухи — спереди назад. Конечно, эмбриологи не могли удержаться и не поиграться с включением-выключением Hox-генов; идеальные для этого объекты — дрозофилы, ведь им от жизни надо немного, они выживают даже при довольно серьезных нарушениях эмбриогенеза. Так вот, если выключить определенный Hox-ген, отвечающий за формирование третьего сегмента груди с последними ножками и жужальцами, то срабатывает другой ген, третий сегмент развивается по образцу второго и получается четырехкрылая муха. Теоретически, если включать гены лишний раз, можно вывести и мушиную многоножку. Если запустить первый Hox-ген и спереди, и сзади, должен получиться дрозофилий Тянитолкай с двумя головами с обеих сторон. Иногда такие нарушения возникают сами собой, и не только с Hox-генами, но и другими, отвечающими за верх-низ, право-лево, конечности, глаза и прочие части тела. Тогда могут рождаться сиамские близнецы, двухголовые, трехрукие и прочие подобные индивиды. Это, ясно, явная патология, но между нормой и патологией грань, как ни странно, очень туманная, так что эволюция подобных регуляторных генов была главным движителем эволюции, ответственным за появление частей тела, отделов черепа и позвоночника, формирование грудной клетки и конечностей, а также всего прочего, чего в протерозое еще не было, но что было вот-вот готово появиться.
Точку в дискуссии поставило открытие Hox-генов — особого семейства генов, определяющих закладку переда, середины и зада. Оказалось, что эти гены мало того что весьма схожи от червей до человека, так они еще и жутко консервативны. Различия же первично- и вторичноротых определяются только разницей в последовательности формирования в общем-то одинаковых частей. Мы начинаем формироваться сзади наперед, а мухи — спереди назад. Конечно, эмбриологи не могли удержаться и не поиграться с включением-выключением Hox-генов; идеальные для этого объекты — дрозофилы, ведь им от жизни надо немного, они выживают даже при довольно серьезных нарушениях эмбриогенеза. Так вот, если выключить определенный Hox-ген, отвечающий за формирование третьего сегмента груди с последними ножками и жужальцами, то срабатывает другой ген, третий сегмент развивается по образцу второго и получается четырехкрылая муха. Теоретически, если включать гены лишний раз, можно вывести и мушиную многоножку. Если запустить первый Hox-ген и спереди, и сзади, должен получиться дрозофилий Тянитолкай с двумя головами с обеих сторон. Иногда такие нарушения возникают сами собой, и не только с Hox-генами, но и другими, отвечающими за верх-низ, право-лево, конечности, глаза и прочие части тела. Тогда могут рождаться сиамские близнецы, двухголовые, трехрукие и прочие подобные индивиды. Это, ясно, явная патология, но между нормой и патологией грань, как ни странно, очень туманная, так что эволюция подобных регуляторных генов была главным движителем эволюции, ответственным за появление частей тела, отделов черепа и позвоночника, формирование грудной клетки и конечностей, а также всего прочего, чего в протерозое еще не было, но что было вот-вот готово появиться.Существуют и другие гены, необычайно похожие у первично- и вторичноротых, до того, что после пересадки гена PAX6 от мыши к мухе чужие гены продолжили работать и дали правильный результат! У мухи с мышиными генами стали появляться дополнительные глаза, причем не мышиные, а фасеточные, ведь гены PAX6 сами не определяют форму и строение органов, а лишь запускают другие нужные в данном месте тела гены.
Некоторое время спустя, в начале кембрия (542 млн лет назад), появились эффективные планктонные фильтраторы, что‑то вроде нынешних рачков. Они ели органическую муть, плавающую в толще воды, и склеивали ее в виде пеллетов, быстро падавших на дно. От этого мутность воды снизилась, а водоросли смогли жить в более глубоких водах. Около дна увеличилось содержание кислорода, так что и эта часть планеты стала доступной аэробам. И тут произошел прорыв!
Увеличение размера клетки ограничено прочностью клеточной мембраны, иначе клетка растечется или даже лопнет. Надо как‑то укреплять границы. Можно нарастить толстую клеточную стенку, как сделали растения и грибы. Но она ограничивает подвижность — трудно бежать, когда ты дерево. Можно нарастить внеклеточную оболочку вокруг всего тела, например хитиновый панцирь членистоногих. Но это в любом случае тяжелая штука, тянущая на дно, где утонувшие задыхались без кислорода. Пока у дна условия были анаэробные и царила тьма, там могли жить лишь чудаковатые хемосинтетики‑анаэробы, а животные и растения не могли стать большими и прочными. Когда же около дна появился кислород, проблема исчезла и перед живыми существами открылись невиданные доселе перспективы. Дальше дна не провалишься. Теперь стало возможным ползать по субстрату, становясь сколь угодно большим и тяжелым. Унылое и безжизненное доселе дно повеселело, расцвело и зашевелилось. Реализация открывшихся возможностей известна как “кембрийская революция”. Собственно, так палеонтологи с геологами и различают слои докембрия и кембрия: в первых мы не видим крупных животных с твердыми покровами, а во вторых их полно. Долгое время “кембрийский взрыв” был загадкой для ученых. Получалось, что мшанки, кораллы, моллюски, брахиоподы, трилобиты и прочая членистоногая нечисть возникли ниоткуда. В реальности их предки жили уже в докембрии, но имели микроскопические размеры и зачаточные твердые покровы.
Долгое время “кембрийский взрыв” был загадкой для ученых. Получалось, что мшанки, кораллы, моллюски, брахиоподы, трилобиты и прочая членистоногая нечисть возникли ниоткуда. В реальности их предки жили уже в докембрии, но имели микроскопические размеры и зачаточные твердые покровы.
В венде — преддверии кембрия — существовали и сравнительно крупные существа, но все они были бесскелетными, а потому их отпечатки сохраняются крайне редко. Самый известный их пример — эдиакарская фауна, сборище фантастических тварей инопланетного облика, лишь малая часть коих имеет родство с современными животными.
Так скажем же спасибо безымянным героям — докембрийским цианобактериям и кембрийским фильтраторам — за наши кости, зубы и ногти, прочность и стойкость, улыбку и прическу, маникюр и педикюр.
Кстати, о морозе…
В конце протерозоя грянуло Лапландское оледенение. Уже одно его название должно нагонять страху на нас, тропических зверей. Но хуже того, оно действительно было самым сильным за всю историю планеты. Земля чуть ли не целиком была скована льдами, а жизнь висела на волоске. Расчеты показывают, что полное покрытие планеты снегом и льдом чревато необратимым остыванием: большая часть доходящего от Солнца тепла будет отражаться обратно в космос, а оставшегося не хватит на растапливание ледников. Если бы такой печальный сценарий реализовался, еще неизвестно, смогли бы выжить хоть какие-то организмы. Наша Земля могла превратиться в аналог Марса.С другой стороны, не исключено, что именно похолодание сыграло положительную роль, так как в холодной воде кислород растворяется лучше; недаром именно арктические и антарктические моря особо богаты жизнью — водорослями, рачками, рыбами и китами.
Полностью читайте:
Дробышевский Станислав, «Достающее звено» (в двух томах). Книга первая. Обезьяны и все-все-все. — М.: Corpus, 2017.
Дробышевский Станислав, «Достающее звено» (в двух томах). Книга вторая. Люди. — М.: Corpus, 2017.
Люди. — М.: Corpus, 2017.
Станислав Дробышевский. Достающее звено
Достающее звено / Станислав Дробышевский. – Москва : АСТ : CORPUS, 2019. Кн. 1 : Обезьяны и все-все-все. – 2019. – 665 с. Кн. 2 : Люди. – 2019. – 588 с.
Вышедший в 2017-м труд Дробышевского был переиздан уже в прошлом году, что лучше любых слов говорит о его популярности. В чём же его секрет?
Прежде всего, «Достающее звено» – очень качественный научпоп. Книга написана легко, понятно и с юмором, но при этом подкреплена солидной научной базой. Даже сейчас, когда после засилья всяческих «Тайн Бермудского треугольника» ниша научно-популярной литературы начала вновь расцветать благодаря работам молодых популяризаторов, это редкое и ценное достоинство. Но есть и другая ось координат: строго научной литературы по антропологии, написанной по-русски и при этом описывающей картину открытий последних десятилетий, описывающей то, как видит историю происхождения человека современная наука, эдакой суммы антропологии – до сих пор не было, точнее, не было очень давно.
В итоге «Достающее звено» заинтересует как читателя, уже подкованного в дисциплине, так и того, кто только собрался разобраться в теории эволюции. А разбираться в ней необыкновенно интересно, да и полезно. И чтобы развеять мифы о ней. И чтобы увидеть историю нашего происхождения с точки зрения науки. И чтобы просто узнать много нового о развитии жизни на Земле, о быте наших предков и отдалённых родственников, о научных методах познания… К тому же, орудия австралопитеков и стоянки кроманьонцев помогают лучше понять современного человека и самого себя. Ведь за сотни тысяч лет некоторые вещи остались неизменными: всё так же альтруизм и способность к обучению, креативность и страсть к познанию «делают человека человеком».
Владислав Романов, библиотекарь Рыбацкой библиотеки
В фонде электронной библиотеке «ЛитРес» аудиокнига.
Книги вас ждут в библиотеках:
- ЦДБ,
- библиотека № 2 им. Ф. Абрамова,
- библиотека № 3 им. О. Ф. Берггольц,
- библиотека № 5 им. Н. Рубцова,
- Рыбацкая библиотека № 6,
- библиотека № 7,
- детская библиотека № 8,
- библиотека № 9 им. Даниила Гранина,
- детская библиотека № 10 им. Н. Носова,
- детская библиотека № 13.
Станислав Дробышевский — Достающее звено. Книга 1. Обезьяны и все-все-все
Станислав Дробышевский
Достающее звено. Книга 1. Обезьяны и все-все-все
© С. Дробышевский, 2017
© Р. Евсеев, иллюстрации, 2017
© Е. Мартыненко, иллюстрации, 2017
© И. Мурашев, иллюстрации, 2017
© О. Федорчук, иллюстрации, 2017
© Д. Хайдаров, иллюстрации, 2017
© А. Бондаренко, оформление, 2017
© ООО “Издательство АСТ”, 2017
Издательство CORPUS ®
* * *Посвящается Инге, Володе и Маше – моей любимой семье
…а также муравьеду и руконожке
Рассказ о цепях коротких и длинных, непрерывных и оборванных, магистральных и параллельных, прямых и извилистых, о том, из каких звеньев они составлены, и о том, все ли звенья достают, как люди достают эти звенья и как звенья достают людей, о звеньях прочных и не очень, о звеньях главных и второстепенных, о звеньях между звеньями и рядом со звеньями, неотвратимости и случайности, о наследии и следах, много о прошлом, в меру о настоящем и немножко о будущем…
Пролог,
в котором автор ведет себя прилично, мило улыбается, много раз говорит спасибо и даже не ехидничает
К написанию этой книги меня сподвигли несколько обстоятельств. Во-первых, преподавательская деятельность, коей я занимаюсь, создает склонность к популярному и доступному объяснению. Во-вторых, большое количество обращений за разъяснениями глобальных или каких-либо частных вопросов антропогенеза со стороны студентов, знакомых и представителей средств массовой информации показало мне, что многим людям эти вопросы небезынтересны. В-третьих, изобилие непрофессиональных, часто просто неграмотных и ошибочных изложений антропогенеза в последнее время зашкаливает: иногда в книгах с заглавием “Антропология” о таковой не говорится вовсе ничего.
Во-вторых, большое количество обращений за разъяснениями глобальных или каких-либо частных вопросов антропогенеза со стороны студентов, знакомых и представителей средств массовой информации показало мне, что многим людям эти вопросы небезынтересны. В-третьих, изобилие непрофессиональных, часто просто неграмотных и ошибочных изложений антропогенеза в последнее время зашкаливает: иногда в книгах с заглавием “Антропология” о таковой не говорится вовсе ничего.
Непосредственным толчком к созданию книги послужило предложение Александра Соколова дать интернет-интервью на тему антропогенеза. На самом деле, уже настойчивостью и терпением в вытряхивании из меня этого интервью Александр сделал огромное дело! Как и подавляющее большинство ученых и преподавателей, я сам мог бы очень долго мечтать о написании такой книги и никогда бы не собраться начать реальные действия. Это вообще дело хлопотное и специфическое. Таких, как я, надо сильно тормошить, и за это Александру огромное спасибо! Первая версия книги опубликована на сайте “Антропогенез.ру” (www.antropogenez.ru ), созданном опять же усилиями в основном Александра. Без этого сайта, даже если бы книга и была когда-нибудь написана, она была бы заметно бледнее и в ней сохранилось бы больше ляпов.
С самого начала интернет-портал “Антропогенез.ру” затевался как популяризаторский, направленный на продвижение знаний в народ и борьбу с лженаучными и околонаучными мифами, с чем Александр успешно справляется. Нельзя не отметить его недавно вышедшую книгу “Мифы об эволюции человека”, уникальную и единственную в своем роде (Соколов, 2015). Ее можно читать параллельно с той, что вы держите в руках, – они не повторяют, а дополняют друг друга.
Образцом научно-просветительского творчества, на который стоит ориентироваться всем популяризаторам, являются книги, лекции и статьи на сайтах www.evolbiol.ru и www.elementy.ru , созданные А. В. Марковым и Е. Б. Наймарк. За это и за важные замечания по содержанию книги им огромное спасибо!
Многие идеи, особенно касающиеся экологических аспектов эволюции, почерпнуты из книг и лекций К. Ю. Еськова, за что я ему бесконечно благодарен.
Ю. Еськова, за что я ему бесконечно благодарен.
Неподражаемый стиль книг Л. Б. Вишняцкого и В. Р. Дольника в немалой степени определил настрой и моего творения. Не всем нравится, когда высоконаучные темы обсуждаются как бы не вполне серьезно, некоторые люди даже искренне считают, что чем ближе фраза к бессмертным образцам “при гулярной ундуляции нет ничего глупее латерального сжатия”, “доминирующая положительная анаболия ведет к девиации” и “вторичную хондрификацию эндоскелета можно рассматривать как гистологическую фетализацию, присущую анамниям”, тем она научнее. Но у меня есть два оправдания. Во-первых, используя человеческую речь вместо “высокой латыни”, я могу надеяться, что труд мой будет-таки осилен до конца большинством читателей, а во-вторых, я сам не засну и не брошу его на полпути. Для любителей подробностей есть приложение в конце книги, тысячи статей и вставленные в книгу “уголки занудства”, для сторонников иных крайностей – “минутки фантазии”, к которым, конечно, не стоит относиться чересчур серьезно.
Отдельное спасибо хочу сказать Дмитрию Владимировичу Богатенкову – моему сокурснику и соратнику в популяризации антропологии. Совместно написанный с ним учебник по антропологии был в свое время издан слишком маленьким тиражом, но его части, посвященные антропогенезу, послужили костяком настоящей книги (кстати, об учебниках: дабы не превращать сию книгу в один из оных, я, скрепя сердце преподавателя, все же проигнорировал просьбы пояснять по ходу текста такие термины, как “цитоплазма” и “АТФ”, – для этого есть школьный курс).
Книга стала, несомненно, лучше после того, как ее критически прочитали и оценили замечательные рецензенты. Я глубоко признателен Светлане Анатольевне Бурлак, благодаря ей текст стал не только корректнее, но и грамотнее. Спасибо Валентине Владимировне Росиной и Ирине Николаевне Грибковой – они взглянули на многие спорные места с неожиданной стороны, отчего удалось прояснить их и, надеюсь, уменьшить число вопросов, которые могли бы задать мне читатели.
И самое большое Спасибо всем читателям нашего портала “Антропогенез.ру” за горячую поддержку и помощь в написании и редактировании этой книги, за вдохновение, которое они в меня вселяют. Без их вопросов, комментариев и советов книга вышла бы гораздо более блеклой, сухой и скучной.
Введение,
в коем возвышенным слогом поясняется, почему книга называется так, как она называется
Эта книга посвящена вроде бы человеку, но речь в ней пойдет далеко не только о нем. Эту странность стоит пояснить. Человек интересен сам себе; следуя незабвенному завету К. Линнея, он усиленно изучает именно себя любимого. Да и странно было бы, если бы основатель систематики глубокомысленно изрек: “Человек, познай выхухоль!” или “Человек, познай пеночку-теньковку!”. Нет, все же речь шла о самом себе. С другой стороны, антропология – биологическая наука, антропологи неизбежно биоцентричны. Антрополог не может мыслить антропоцентрично по определению, как бы странно это ни звучало. Человек – неотделимая часть биологического мира, понять его можно, только разобравшись с иными живыми существами и взаимосвязями между ними.
Начиная с первой публикации книги Ч. Дарвина “Происхождение видов путем естественного отбора” (несмотря на то, что в ней ничего прямо не говорилось об эволюции человека), среди естествоиспытателей пошли споры о “недостающем звене” – промежуточной форме между обезьяной и человеком. Особенно активно пропагандировал существование этого звена Э. Геккель, предположивший существование в прошлом вида Pithecanthropus alalus – “обезьяночеловека бессловесного”, название которого частично использовал Э. Дюбуа в 1894 году для описания Pithecanthropus erectus – “обезьяночеловека прямоходящего”. Однако Э. Геккель своего “обезьяночеловека” придумал, а Э. Дюбуа – нашел на острове Ява, в виде черепной крышки и бедренной кости.
Сначала казалось, что картина эволюции человека окончательно прояснилась, тем более что позднейшие ступени тоже представлялись известными, ведь еще в 1857 году немецкий учитель К. Фульрот явил миру найденного годом ранее неандертальца – как многие тогда считали, прямого предшественника современного человека, не имевшего уже почти никаких обезьяньих черт. Цепочка “обезьяна – питекантроп – неандерталец – сапиенс” выглядела вполне достаточной и полной для окончательного торжества научной точки зрения над ортодоксальной религиозной (характерно, что современные школьные учебники недалеко ушли от воззрений конца XIX века). Однако сомнения никогда не оставляют человеческий разум: тогда как К. Майер доказывал, что скелет из Неандерталя является останками русского казака-дегенерата, умершего в пещере от тягот погони за Наполеоном (слишком покатый лоб он объяснял врожденной патологией, а слишком кривые ноги – постоянной верховой ездой), Э. Краузе обосновывал принадлежность костей с Явы гигантскому гиббону (объясняя этим опять же слишком покатый лоб и “слишком” прямую бедренную кость). Новые находки развеяли этот туман, но загадали новые загадки. В первые десятилетия XX века питекантроп перестал казаться таким уж обезьяноподобным, но тут встал закономерный вопрос: а где же в таком случае “недостающее звено” между питекантропом и обезьяной? С этих пор погоня за “недостающим звеном” уже не прекращалась. Не помогло ни описание Р. Дартом австралопитека из Южной Африки, ни открытие “с другого конца” эволюционной линии – африканских проконсулов. Весь XX век разрыв между “обезьяньим” и “человеческим” концами сокращался, но сближение это напоминает погоню Ахиллеса за черепахой в апории Зенона – всегда кажется недостаточным, неполным, незавершенным.
Фульрот явил миру найденного годом ранее неандертальца – как многие тогда считали, прямого предшественника современного человека, не имевшего уже почти никаких обезьяньих черт. Цепочка “обезьяна – питекантроп – неандерталец – сапиенс” выглядела вполне достаточной и полной для окончательного торжества научной точки зрения над ортодоксальной религиозной (характерно, что современные школьные учебники недалеко ушли от воззрений конца XIX века). Однако сомнения никогда не оставляют человеческий разум: тогда как К. Майер доказывал, что скелет из Неандерталя является останками русского казака-дегенерата, умершего в пещере от тягот погони за Наполеоном (слишком покатый лоб он объяснял врожденной патологией, а слишком кривые ноги – постоянной верховой ездой), Э. Краузе обосновывал принадлежность костей с Явы гигантскому гиббону (объясняя этим опять же слишком покатый лоб и “слишком” прямую бедренную кость). Новые находки развеяли этот туман, но загадали новые загадки. В первые десятилетия XX века питекантроп перестал казаться таким уж обезьяноподобным, но тут встал закономерный вопрос: а где же в таком случае “недостающее звено” между питекантропом и обезьяной? С этих пор погоня за “недостающим звеном” уже не прекращалась. Не помогло ни описание Р. Дартом австралопитека из Южной Африки, ни открытие “с другого конца” эволюционной линии – африканских проконсулов. Весь XX век разрыв между “обезьяньим” и “человеческим” концами сокращался, но сближение это напоминает погоню Ахиллеса за черепахой в апории Зенона – всегда кажется недостаточным, неполным, незавершенным.
ПОНРАВИЛАСЬ КНИГА?
Эта книга стоит меньше чем чашка кофе!
УЗНАТЬ ЦЕНУ
Станислав Дробышевский: «Достающее звено». Рецензия
Двухтомник Станислава Дробышевского «Достающее звено» — находка для желающих узнать об антропогенезе с самого его начала и до настоящего времени. Но подойдёт она не каждому — для продуктивного чтения нужно быть хотя бы немного подкованным в эволюции человека и биологии вообще. Автор порой с непринуждённой лёгкостью и шутками, а порой уходя в сложную для понимания узкоспециальную терминологию и начиная перечислять множество латинских названий, повествует об антропогенезе. Эти переходы от одного стиля к другому затрудняют восприятие прочитанного. Антропогенез в книге рассматривается на довольно протяжённом интервале: здесь есть сведения и о появлении жизни на Земле, и о «переходных звеньях», и даже совсем кратко обсуждается предполагаемое будущее человечества. Помимо биологической грани эволюции человека, автор рассказывает об эволюции живого мира вообще, антропологии, генетике, культурных революциях и их влиянии на антропогенез. Именно поэтому у читателя формируется наиболее полное понимание этого процесса, в который вовлечено множество зависящих друг от друга факторов и событий.
Автор порой с непринуждённой лёгкостью и шутками, а порой уходя в сложную для понимания узкоспециальную терминологию и начиная перечислять множество латинских названий, повествует об антропогенезе. Эти переходы от одного стиля к другому затрудняют восприятие прочитанного. Антропогенез в книге рассматривается на довольно протяжённом интервале: здесь есть сведения и о появлении жизни на Земле, и о «переходных звеньях», и даже совсем кратко обсуждается предполагаемое будущее человечества. Помимо биологической грани эволюции человека, автор рассказывает об эволюции живого мира вообще, антропологии, генетике, культурных революциях и их влиянии на антропогенез. Именно поэтому у читателя формируется наиболее полное понимание этого процесса, в который вовлечено множество зависящих друг от друга факторов и событий.
…вопрос о «недостающем звене» уже более полувека не стоит в научной повестке дня; сейчас известны и подробно описаны практически все переходы от непосредственных предков приматов до современного человека.
Станислав Дробышевский, «Достающее звено»
В своём двухтомнике «Достающее звено» Станислав Дробышевский рассказывает читателю об антропогенезе, освещая эту тему с различных сторон — он рассуждает и с биологической, и с археологический, и с культурологической точки зрения. В этом книга немного напоминает «Sapiens: Краткая история человечества» Юваля Ной Харари.
Станислав Дробышевский — известный российский антрополог, в библиографии которого немало работ об эволюции человека, в том числе и научно-популярных. В «Достающем звене», он штрих за штрихом создаёт у читателя понимание того, какие предпосылки и события способствовали появлению предков человека, предков предков человека и так далее, вплоть до появления жизни на Земле. В первой книге речь идёт о животных, преимущественно обезьянах, и об их признаках, которые похожи (или непохожи) на таковые у наших предков. Автор приводит много сведений из зоологии, эволюции, и не только для человека, но и для других организмов. Например, он прослеживает, как менялся конкретный признак у разных живых существ по мере эволюции животного мира, попутно рассказывая о рыбах, миксинах и ланцетниках. А во второй книге Станислав Дробышевский повествует уже об эволюции человека, начиная с сахелянтропа и заканчивая собственно современным человеком и, более того, альтернативными «звеньями» из его будущего с примерами морфологических, поведенческих и культурных изменений.
Например, он прослеживает, как менялся конкретный признак у разных живых существ по мере эволюции животного мира, попутно рассказывая о рыбах, миксинах и ланцетниках. А во второй книге Станислав Дробышевский повествует уже об эволюции человека, начиная с сахелянтропа и заканчивая собственно современным человеком и, более того, альтернативными «звеньями» из его будущего с примерами морфологических, поведенческих и культурных изменений.
Известно, что добывание мяса требует больших интеллектуальных усилий, чем растительноядное существование. Трава не сопротивляется и не прячется, а мясо обычно не хочет, чтобы его съели, — убегает, лягается и всячески активно избегает хищника.
Станислав Дробышевский, «Достающее звено»
Такой большой объём информации и протяжённый по времени период, который пытается охватить автор, сложно описать простым и понятным языком. Поэтому в тексте часто присутствуют неожиданные смены стиля повествования, когда вначале автор рассказывает о забавном и весёлом, а затем начинает использовать научную терминологию и латынь, словно в серьёзной монографии. Например, он легко и с юмором пишет о хвосте и его роли в жизни рыбообразных, и тут же в подробностях и с терминами говорит об эволюции жаберных дуг. Это периодически выбивает из колеи, и приходится вновь находить нить повествования, искать непонятные термины в других источниках и перечитывать текст ещё раз. Помогает то, что вещи, сложные для понимания неспециалиста, автор нередко выносит во врезки под названием «Уголок занудства». Поэтому к смене серьёзной речи на лёгкую и обратно постепенно привыкаешь, а некоторые моменты, которые для читателя непонятны, сложны или просто пока неинтересны, можно пропустить, а затем, при желании, вернуться к ним вновь. Всё это дополняется большим числом чёрно-белых иллюстраций в электронной версии, а в печатной есть ещё и цветная вкладка. Они служат хорошим подспорьем для понимания материала. А уж совсем заинтересованного в деталях и подробностях читателя в конце каждой книги ждёт обширный список использованной литературы
Если бы я написал классический учебник по антропологии, он был бы короче и информативнее, но шансов на прочтение у него было бы меньше, чем у популярной книжки, текст которой разбавлен всяческими хохмами.
Станислав Дробышевский, «Достающее звено»

Двухтомник «Достающее звено» может быть интересен разношёрстой, но подготовленной аудитории, уже обладающей знаниями в биологии и антропологии: и студентам, и непосредственно учёным, специализирующимся в естественно-научных дисциплинах, и другим подготовленным читателям. Наверняка и школьник сможет найти для себя интересное и доступное для понимания. Но чтение потребует сосредоточенности, чтобы внимательно следить за подчас сложно написанным текстом и за сменой стилей повествования, которые присутствуют в книге. Несмотря на это замечание, «Достающее звено» — многогранный труд, который во время чтения порой вызывает улыбку от юмора автора, а порой заставляет задуматься над сложным материалом и пройтись по перечню ссылок к главе. И именно из-за этого здесь всегда можно будет найти для себя что-то интересное.
Дробышевский А., Бауманн С.Б., Шнайдер В. Батарея быстрых задач фМРТ для картирования зрительных, моторных, когнитивных и эмоциональных функций. Neuroimage 31: 732-744
Позитронно-эмиссионная томография (ПЭТ) и функциональная магнитно-резонансная томография (фМРТ) широко используются для исследования функциональной нейроанатомии когнитивных функций. Здесь мы рассматриваем 275 исследований ПЭТ и фМРТ внимания (устойчивое, выборочное, Stroop, ориентация, разделенное), восприятия (объект, лицо, пространство / движение, запах), образов (объект, пространство / движение), языка (письменное / устное слово). распознавание, речевой / неречевой ответ), рабочая память (вербальная / числовая, объектная, пространственная, для решения проблем), семантическое извлечение памяти (категоризация, генерация), эпизодическое кодирование памяти (вербальное, объектное, пространственное), эпизодическое извлечение памяти (вербальное, невербальная, успех, усилие, режим, контекст), прайминг (перцептивная, концептуальная) и процедурная память (кондиционирование, моторные и немоторные навыки). Чтобы определить последовательные паттерны активации, связанные с этими когнитивными операциями, данные из 412 контрастов были обобщены на уровне кортикальных областей Бродмана, островка, таламуса, медиально-височной доли (включая гиппокамп), базальных ганглиев и мозжечка. Что касается восприятия и образов, паттерны активации включали первичные и вторичные области дорсальных и вентральных путей. Что касается внимания и рабочей памяти, активация обычно обнаруживалась в префронтальной и теменной областях. Для восстановления языка и семантической памяти типичные области включали левую префронтальную и височную области.Для кодирования эпизодической памяти последовательно активированные области включали левую префронтальную и медиальную височные области. Для восстановления эпизодической памяти образцы активации включали префронтальную, медиальную височную и заднюю срединные области. Для прайминга постоянно наблюдались деактивации в префронтальной (концептуальной) или экстрастриарной (перцептивной) областях. Что касается процедурной памяти, активация была обнаружена как в моторных, так и в немоторных областях мозга. Анализ региональных активаций в когнитивных доменах показал, что несколько областей мозга, включая мозжечок, задействованы в различных когнитивных проблемах.Эти наблюдения обсуждаются в отношении функциональной специализации, а также функциональной интеграции.
Чтобы определить последовательные паттерны активации, связанные с этими когнитивными операциями, данные из 412 контрастов были обобщены на уровне кортикальных областей Бродмана, островка, таламуса, медиально-височной доли (включая гиппокамп), базальных ганглиев и мозжечка. Что касается восприятия и образов, паттерны активации включали первичные и вторичные области дорсальных и вентральных путей. Что касается внимания и рабочей памяти, активация обычно обнаруживалась в префронтальной и теменной областях. Для восстановления языка и семантической памяти типичные области включали левую префронтальную и височную области.Для кодирования эпизодической памяти последовательно активированные области включали левую префронтальную и медиальную височные области. Для восстановления эпизодической памяти образцы активации включали префронтальную, медиальную височную и заднюю срединные области. Для прайминга постоянно наблюдались деактивации в префронтальной (концептуальной) или экстрастриарной (перцептивной) областях. Что касается процедурной памяти, активация была обнаружена как в моторных, так и в немоторных областях мозга. Анализ региональных активаций в когнитивных доменах показал, что несколько областей мозга, включая мозжечок, задействованы в различных когнитивных проблемах.Эти наблюдения обсуждаются в отношении функциональной специализации, а также функциональной интеграции.
Антропологические характеристики некоторых групп коренных народов в современной Индонезии
Belwood, P., P История индо-малайзийского архипелага , Гонолулу: Univ. Hawaii Press, 2007.
Google ученый
Чуан Т.К., Хартоно М. и Кумар Н., Антропология населения Сингапура и Индонезии, Int.J. Ind. Ergon. , 2010, т. 40, нет. 6. С. 757–766.
Артикул Google ученый
 org/Book»> 3.
org/Book»> 3.Ивановский А.А., Население земного шара. Опыт антропологической классификации , Москва: Тип. П. Рябушинскаго, 1911.
Google ученый
Brown, C., Краткая история Индонезии: маловероятная нация? , Сингапур: Talisman Publ.Pte Ltd., 2011.
Google ученый
Wallace, A.R., The Malay Archipelago , Singapore: Periplus Ed., 2008.
Google ученый
Дерябин В.Е., Построение типологии пропорций тела методом главных компонент // Проблемы эволюционной морфологии человека и его расы , Алексеев В.П. и Зубов А.А., ред., М .: Наука, 1986.
,Крогман, В.М., Физическая антропология индейцев-семинолов Оклахомы, Итальянцев из Комитато за Studie dei Problemi della Popolazione , 1934, т. 2, вып. 3, стр. 119.
Google ученый
Коганей Ю., Beitrage zur physischen Anthropologie der Aino. I. Untersuchungen am Skelett, Mitt. Med. Fachsch. Кайс. Univ. Токио , 1893 г., т.2. С. 1–249.
Google ученый
Такаяма, Х., Соматометрическое исследование Нисеи (японских иммигрантов 2-го поколения), Лима, Перу, Антрополь. Sci. , 1998, т. 106, доп. С. 189–201.
Артикул Google ученый
 org/ScholarlyArticle»> 10.
org/ScholarlyArticle»> 10.Курису К., Многомерный статистический анализ физических взаимоотношений коренных племен в Сараваке, Малайзия, Am.J. Phys. Антрополь. , 1970, т. 33, нет. 2. С. 229–234.
CAS Статья PubMed Google ученый
Лин, И-Ч., Ван, М.-Дж., И Ван, Э. М., Сравнение антропометрических характеристик у четырех народов Восточной Азии, Appl. Эргон. , 2004, т. 35, нет. 2. С. 173–178.
Артикул PubMed Google ученый
Прокопец, М., Антропометрическое исследование Рембарранги: сравнение с другими популяциями, J. Hum. Evol. , 1977, т. 6, вып. 4. С. 371–391.
Артикул Google ученый
Сузуки Х. и Кучи М., Соматометрические данные китайцев, J. Anthropol. Soc. Nippon , 1986, т. 94, нет. 2. С. 177–181.
Артикул Google ученый
Чжэн Л., Ли, Ю., Лу, С., Бао, Дж., Ван, Дж., Чжан, X., Сюэ, Х., Ронг, В., Физические характеристики китайского хакка, Sci. Китай: Life Sci. , 2013, т. 56, нет. 6. С. 541–551.
Артикул Google ученый
Уэллс, Дж. К. К., Экогеографические ассоциации между климатом и составом человеческого тела: анализ на основе антропометрии и кожных складок, Am. J. Phys. Антрополь. , 2012, т. 147, нет. 2. С. 169–186.
Артикул PubMed Google ученый
 org/ScholarlyArticle»> 16.
org/ScholarlyArticle»> 16.Тилкенс, М.Дж., Уолл-Шеффлер, С., Уивер, Т., и Штудель-Числа, К., Влияние пропорций тела на терморегуляцию: экспериментальная оценка правила Аллена, J. Hum. Evol. , 2007, т. 53, нет. 3. С. 286–291.
Артикул Google ученый
В чем разница между кодами состояния HTTP 301 и 308?
Обзор
301 , 302 и 307 RFC 7231, текущий справочник по семантике и содержанию HTTP / 1.1, определяет код состояния 301 (перемещен навсегда) и 302 (найден), что позволяет изменить метод запроса с POST на GET . Эта спецификация также определяет код состояния 307 (временное перенаправление), который не позволяет изменять метод запроса с POST на GET .
Подробнее см. Ниже:
6.4.2. 301 Перемещено навсегда
Код состояния
301(перемещен навсегда) указывает, что целевой объект ресурсу был назначен новый постоянный URI и все будущие ссылки на этот ресурс должны использовать один из вложенных URI.[…]Примечание: По историческим причинам пользовательский агент МОЖЕТ изменить запрос. метод от
POSTдоGETдля последующего запроса. Если это поведение нежелательно, код состояния307(временное перенаправление) можно использовать вместо этого.
6.4.3. 302 Найдено
Код состояния
302(Найдено) указывает, что целевой ресурс временно находится под другим URI.Поскольку перенаправление может быть изменен при случае, клиент должен продолжать использовать действующий URI запроса для будущих запросов. […]Примечание: По историческим причинам пользовательский агент МОЖЕТ изменить запрос.
метод от
POSTдоGETдля последующего запроса. Если это поведение нежелательно, код состояния307(временное перенаправление) можно использовать вместо этого.
6.4.7. 307 Временное перенаправление
Код состояния
307(временное перенаправление) указывает, что целевой ресурс временно находится под другим URI и пользовательским агентом НЕ ДОЛЖЕН изменять метод запроса, если он выполняет автоматическое перенаправление на этот URI.Поскольку перенаправление может меняться со временем, клиент должен продолжать использовать исходный действующий URI запроса для будущих запросов. […]Примечание: Этот код состояния аналогичен
302(Найдено), за исключением того, что он не позволяет изменить метод запроса сPOSTнаGET. Этот Спецификация не определяет эквивалентного аналога для301(перемещено Постоянно) (RFC 7238, однако, определяет код состояния308(Постоянное перенаправление) для этой цели).
Потребность
308 RFC 7238 был создан для определения кода состояния 308 (постоянное перенаправление), который аналогичен 301 (перемещен навсегда), но не позволяет изменять метод запроса с POST на GET .
Код состояния 308 теперь определяется RFC 7538 (который устарел RFC 7238).
3. 308 Постоянное перенаправление
Код состояния
308(постоянное перенаправление) указывает, что целевой ресурсу был назначен новый постоянный URI и все будущие ссылки на этот ресурс должны использовать один из вложенных URI.Клиенты с возможностью редактирования ссылок должны автоматически повторно ссылаться ссылки на действующий URI запроса к одна или несколько новых ссылок, отправленных сервером, где это возможно.[…]
Примечание: Этот код состояния аналогичен
301(перемещен навсегда), за исключением того, что он не позволяет изменить метод запроса сPOSTдоGET.
Se имеем следующее:
+ ----------- + ----------- +
| Постоянный | Временный |
+ ------------------------------------------------- ----------- + ----------- + ----------- +
| Позволяет изменить метод запроса с POST на GET | 301 | 302 |
+ ------------------------------------------------- ----------- + ----------- + ----------- +
| Не позволяет изменить метод запроса с POST на GET | 308 | 307 |
+ ------------------------------------------------- ----------- + ----------- + ----------- +
Выбор наиболее подходящего кода статуса
Майкл Кропат составил набор диаграмм решений, которые помогают определить лучший код статуса для каждой ситуации.См. Следующие коды состояния 2xx и 3xx :
Влияние воздействия изофлурановой анестезии новорожденных на обучающую и сенсорную системы у взрослых
Объекты и обоснование времени
76% детей получают однократное воздействие анестезии, тогда как 24% подвергаются многократному воздействию 51 . 61% этих воздействий имеют продолжительность менее 1,5 часа, а оставшиеся 39% — 1,5 часа или более 51 .Таким образом, для этого исследования были использованы три отдельных воздействия продолжительностью 2 часа каждое на 8, 11 и 14 дни для представления тяжелых случаев на основании вывода о том, что дети, подвергшиеся многократному двухчасовому воздействию анестезии (из-за нескольких операций и / или МРТ 52 ) продемонстрировал особенно высокий риск развития нарушений обучаемости 51,53 . Развитие мозга кролика, в том числе кортикальной ГАМКергической системы, происходит в основном в перинатальном периоде 54 , как и у человека, в отличие от других видов, таких как грызуны, или более ранних видов, таких как обезьяны и овцы. Первая временная точка в 8 дней была выбрана для новорожденных кроликов, поскольку было показано, что обширная миелинизация начинается с 8 дней после рождения 55 .
Первая временная точка в 8 дней была выбрана для новорожденных кроликов, поскольку было показано, что обширная миелинизация начинается с 8 дней после рождения 55 .
Препараты для животных
Кроликов с голландским поясом (N = 18) использовали в соответствии с руководящими принципами Национального института здравоохранения и протоколом, утвержденным Комитетом по уходу и использованию институционального института системы здравоохранения Университета Северного Шора.
Для подготовки животных мы следовали ранее опубликованному протоколу 13 .Наборы кроликов родились в гнездовом ящике, содержащем измельченную подстилку из осины, которая была приготовлена заранее. Наборы новорожденных кроликов содержались и содержались вместе с матерью до 4–6 недель, что является оптимальным возрастом отъема для голландских кроликов. Наборы от каждой плотины были случайным образом распределены по анестезиологической и контрольной группам. Начиная с 8-го дня после рождения, наборы в группе анестезии анестезировали индивидуально с помощью маски для носа. Наборы в контрольной группе подвергались воздействию той же среды, что и группа анестезии, при дыхании воздухом.Изофлуран вводили с использованием испарителей Vapor 19.1 в течение 2 часов при 1 ПДК (~ 2%) в воздухе или в 80% -ном кислороде с канистрой для удаления на стороне выпуска. Концентрация 1MAC была подтверждена с помощью техники зажима за хвост. Кроликов подвергали воздействию анестезии на 8, 11 и 14 дни по 2 часа каждое воздействие. Во время анестезии каждые 15 мин контролировали температуру тела и дыхание. Грелка с рециркуляцией теплой воды (T-Pump, Gaymar Industries Inc, Orchard Park, NY, USA) использовалась для контроля температуры.Контрольную группу помещали в тот же ящик на тот же срок, что и группу анестезии. После каждого сеанса кроликов возвращали в плотину. Были предприняты все усилия, чтобы предотвратить отторжение кроликов плотиной (например, комплекты были завернуты в материал для гнезд, чтобы поддерживать нормальный запах). Ни один из комплектов не был отвергнут плотиной. Всего было использовано 18 кроликов. Предварительные поведенческие данные и данные фМРТ от 10 кроликов в контрольной группе и группе с анестезией в воздухе (IA) были представлены ранее и были проанализированы вместе с дополнительными 8 кроликами для настоящего исследования.
Ни один из комплектов не был отвергнут плотиной. Всего было использовано 18 кроликов. Предварительные поведенческие данные и данные фМРТ от 10 кроликов в контрольной группе и группе с анестезией в воздухе (IA) были представлены ранее и были проанализированы вместе с дополнительными 8 кроликами для настоящего исследования.
В возрасте 3 месяцев и самкам, и самцам были имплантированы фиксирующие головные болты, и они привыкли к среде МРТ, как описано ранее 56 . Животных анестезировали смесью кетамина (60 мг / кг) и ксилазина (10 мг / кг). В черепе было проделано 5–6 мелких боровых отверстий без полного проникновения кости. Маленькие нейлоновые поддерживающие винты вставлялись в бортовые отверстия. Легкое подголовник, содержащее четыре нейлоновых болта, было имплантировано наверху черепа.Это устройство также служило для позиционирования ВЧ-катушки в стереотаксической плоскости во время МР-экспериментов. Рана была полностью покрыта зубным цементом, при необходимости накладывались нерассасывающиеся швы. После 1 недели восстановления после операции каждый субъект привык в течение 3-5 дней к среде визуализации перед экспериментами. 2 кролика получили анестезию, но были исключены из исследований, потому что их имплант был сломан. Окончательное количество кроликов в каждой группе для функциональных исследований in vivo составляло: N = 6 (3 самки и 3 самца) для контроля, N = 6 (4 самки и 2 самца) для IA и N = 4 (1 самка и 3 самца). мужчины) для анестезии кислородом (IO).
Классическое кондиционирование Eyeblink
ECC — это хорошо контролируемый тест, обычно используемый как у животных, так и у людей, в котором субъект учится связывать нейтральный условный раздражитель (CS) с поведенческим безусловным стимулом (US). Во время экспериментов кроликам выдавали беруши, чтобы уменьшить потенциально отвлекающий шум из окружающей среды. CS был доставлен путем отклонения двух усов (A1 и B1), прикрепленных к волоконной ленте на левой стороне кролика, с амплитудой 1. 5 мм и частотой 50 Гц, используя систему, описанную ранее 57 . США состояли из воздушного потока 3 фунта на квадратный дюйм, который подавался сжатым воздухом и управлялся регулятором и соленоидным клапаном. УЗИ поставили на левый глаз. Движение век измеряли с помощью оптоволоконного инфракрасного датчика отражения 58 , который располагался на расстоянии 1 см от роговицы. Длительность CS и US составляла 250 и 150 мс, соответственно, с интервалом следа без стимула 500 мс. Общая продолжительность одного испытания составляла 8 с, со случайным интервалом между испытаниями 5–10 с.CR был определен как изменение напряжения детектора, которое было на 4 SD больше, чем средняя базовая амплитуда и произошло по крайней мере через 35 мс после начала CS, но до US. Данные Eyeblink были взяты с частотой 300 Гц. Каждый субъект проходил 1 сеанс кондиционирования в день в течение 10 дней, при этом каждый сеанс состоял из 100 попыток. Животных обучали внутри магнита, используемого для фМРТ, без присутствия шума от импульсных градиентов поля. После тренировки на 10-й день животные прошли 20 дополнительных испытаний без прикрепления усов к волокнистой ленте, чтобы подтвердить, что испытуемые были подготовлены только к стимуляции усов, а не к звуку вибрации.Ни один из кроликов не показал более 5% CR в этих испытаниях.
5 мм и частотой 50 Гц, используя систему, описанную ранее 57 . США состояли из воздушного потока 3 фунта на квадратный дюйм, который подавался сжатым воздухом и управлялся регулятором и соленоидным клапаном. УЗИ поставили на левый глаз. Движение век измеряли с помощью оптоволоконного инфракрасного датчика отражения 58 , который располагался на расстоянии 1 см от роговицы. Длительность CS и US составляла 250 и 150 мс, соответственно, с интервалом следа без стимула 500 мс. Общая продолжительность одного испытания составляла 8 с, со случайным интервалом между испытаниями 5–10 с.CR был определен как изменение напряжения детектора, которое было на 4 SD больше, чем средняя базовая амплитуда и произошло по крайней мере через 35 мс после начала CS, но до US. Данные Eyeblink были взяты с частотой 300 Гц. Каждый субъект проходил 1 сеанс кондиционирования в день в течение 10 дней, при этом каждый сеанс состоял из 100 попыток. Животных обучали внутри магнита, используемого для фМРТ, без присутствия шума от импульсных градиентов поля. После тренировки на 10-й день животные прошли 20 дополнительных испытаний без прикрепления усов к волокнистой ленте, чтобы подтвердить, что испытуемые были подготовлены только к стимуляции усов, а не к звуку вибрации.Ни один из кроликов не показал более 5% CR в этих испытаниях.
Доставка стимула фМРТ и сбор данных
В каждом эксперименте данные фМРТ получали от бодрствующих кроликов в ответ на стимуляцию усов. Стимуляция усов осуществлялась путем отклонения двух усов (A1 и B1) с амплитудой 1,5 мм и частотой 50 Гц с использованием системы, описанной ранее 57 . Каждое животное получало 10 попыток стимуляции усов, которые состояли из базового (30 с), стимуляционного (20 с) и постстимулирующего (40 с) периодов до и после тренировки с парадигмой ECC.
эксперименты с фМРТ проводили, как описано ранее 13 , с использованием спектрометра визуализации 9,4 Т (BioSpec 94 / 30USR, Bruker Biospin MRI GmbH), работающего на частоте 1H 400 МГц. Спектрометр был снабжен активно экранированной градиентной катушкой (BFG-240–150-S-7, Research Resonance, Inc., Биллерика, Массачусетс, США). Как для передачи, так и для приема использовалась одновитковая круговая поверхностная ВЧ-катушка диаметром 40 мм. Перед каждым экспериментом были получены анатомические изображения с использованием последовательности импульсов градиентного эхо-сигнала с множеством срезов с TR, равным 1.5 с, TE 10 мс, поле зрения 30 мм × 30 мм и размер матрицы 128 × 128, что соответствует разрешению в плоскости 234 мкм × 234 мкм. Данные фМРТ были получены из четырех последовательных в аксиальной плоскости с использованием одноразовой многосрезовой последовательности градиентного эхо-сигнала EPI с временем повторения (TR) 2 с, временем эхо-сигнала (TE) 11 мс, 30 мм × Поле зрения 30 мм (FOV) и размер матрицы 80 × 80, что соответствует размеру вокселя в плоскости 375 мкм × 375 мкм и толщине среза 1 мм.
Данные фМРТ были скорректированы на небольшое движение головы с использованием метода аффинной двумерной регистрации, реализованного с использованием инструментария Insight ITK 59 , а последующая обработка была реализована в Matlab (The MathWorks, Inc., Натик, Массачусетс). Перед усреднением испытания были проверены на наличие любых оставшихся движений головы. Затем испытания были усреднены, и активированные воксели были обнаружены в усредненных данных с использованием алгоритма на основе неконтролируемой одноклассной машины опорных векторов (SVM), как описано ранее 60 .
ЖИРНЫЕ временные отклики были получены путем усреднения активированных вокселей для каждой области. Чтобы сравнить временные реакции более подробно, были определены начальная и вторичная фазы.Первоначальный ответ был определен как максимум в течение первых пяти временных точек стимуляции. Вторичный ответ определялся как максимум за последние пять точек стимуляции 29 . Чтобы сравнить максимумы ЖИРНЫХ ответов, временные курсы были нормализованы к среднему значению первых пяти временных точек стимуляции, чтобы подавить индивидуальные различия в ЖИРНОЙ величине.
Объем гиппокампа
Объемы левого и правого полушарий дорсального гиппокампа были измерены по анатомическим изображениям, полученным до (3 месяца после последнего воздействия анестезии) и после обучения (3.5 месяцев после последнего воздействия анестезии). Изображения были получены с использованием последовательности импульсов градиентного эхо-сигнала с несколькими срезами с TR 1,5 с, TE 10 мс, 30 мм × 30 мм FOV и размером матрицы 128 × 128, что соответствует разрешению в плоскости 234 мкм × 234 мкм. Объем гиппокампа измеряли на одном и том же срезе, содержащем дорсальный гиппокамп, в оба момента времени (до и после обучения). Мозг был тщательно позиционирован с использованием известных анатомических ориентиров, чтобы представить один и тот же срез у отдельных животных. Гиппокамп был вручную очерчен в Bruker Paravision 5.1 и объемы левого и правого гиппокампа, нормализованные к объему всего среза для каждого животного.
Протонная МР-спектроскопия
Бодрствующих кроликов сканировали на спектрометре визуализации Bruker Biospec 9.4T. Изображения курсового радиомаяка были получены с использованием T 2 -взвешенной последовательности RARE-импульсов. Спектры протонов с подавлением воды были получены из изотропных 2-миллиметровых вокселей, расположенных в левой и правой зубчатой извилине, с использованием локализации PRESS и подавления воды VAPOR. Использовались TR / TE 2500 мс / 10 мс, спектральная ширина 4000 Гц, 8192 комплексных точки и 256 средних значений.Однородность магнитного поля была оптимизирована для каждого из выбранных вокселей с использованием метода быстрой карты до получения спектра из этого вокселя. NAA, глутамат, глутамин, ГАМК, креатин, холин и мио-инозит были количественно определены с использованием анализа модели ЖХ, и их уровни выражены как отношение к общему креатину в том же спектре. Количество кроликов, исследованных MRS для каждой группы, следующее: N = 5 (3 самки и 2 самца) для контроля (1 кролик сломал головной болт перед этим типом визуализации и был умерщвлен), N = 6 для IA (4 самки и 2 самца) и N = 4 для ИО (1 самка и 3 самца).
Визуализация тензора диффузии
Измерения DTI проводили ex vivo на фиксированных препаратах головного мозга. Кроликов в возрасте 4,5 месяцев глубоко анестезировали кетамином и ксилазином и подвергали эвтаназии путем перфузии 0,9% физиологическим раствором с последующим введением 10% формалина в фосфатно-солевом буфере через левую внутрижелудочковую инъекцию. Затем мозг был зафиксирован в формалине. Всего 9 мозгов (4 из контрольной группы (2 женщины и 2 мужчины) и 5 из группы анестезии, состоящей из субъектов IA и IO (3 женщины и 2 мужчины) были исследованы с помощью DTI, а другие мозги были выброшены из-за повреждений во время подготовки.
Для DTI мозг регидратировали PBS в течение 48 часов для удаления фиксатора, а затем помещали в пробирку для образца и окружали фомблином, чтобы предотвратить высыхание и минимизировать градиенты чувствительности между мозгом и окружающей средой.
DTI-данные были получены на формирователе изображения 9,4 Т в аксиальной плоскости с помощью протокола многосрезового спин-эхо-визуализации, взвешенного по 64 направлениям, с использованием следующих параметров: TR = 4000 мс, TE = 24,9 мс, na = 1, один b Значение = 1000 с / мм 2 , длительность градиента диффузии δ = 7 мс, разделение градиентов Δ = 11.2 мс, два изображения A0, количество срезов 16, толщина среза 0,5 мм, FOV 34,5 мм × 45,5 мм, матрица 256 × 192 для разрешения в плоскости 135 мкм x 237 мкм. Изображения были обработаны с использованием программного обеспечения Paravision 5.1 для создания карт фракционной анизотропии (FA) и коэффициента диффузии по осям x, y и z. Следующие интересующие области были вручную очерчены как на левом, так и на правом полушариях мозга на изображениях параметров FA: дорсальный гиппокамп CA1 и DG (дорсальная и вентральная лопатки), фимбрия, мозолистое тело, лучистая корона, внутренняя и внешняя капсула.Осевой коэффициент диффузии (AD) и радиальный коэффициент диффузии (RD) рассчитывали из измеренных значений коэффициентов диффузии в направлениях x, y и z. Значения FA, AD и RD для обоих полушарий усреднялись и сравнивались между контрольной группой и группой анестезии. Поскольку их показатели распространения были почти идентичны, группы IA и IO были объединены для анализа, чтобы увеличить статистическую мощность.
Значения FA, AD и RD для обоих полушарий усреднялись и сравнивались между контрольной группой и группой анестезии. Поскольку их показатели распространения были почти идентичны, группы IA и IO были объединены для анализа, чтобы увеличить статистическую мощность.
Статистический анализ
Парный t-критерий использовался для сравнения пар «до — после обучения» или «левая — правая сторона» у одних и тех же животных.Факторный дисперсионный анализ ANOVA (Statistica, StatSoft, Tulsa, OK) использовался для определения значимости разницы между средними значениями в группах, подвергнутых анестезии, и в контрольных группах. Факторный дисперсионный анализ ANOVA использовался с 3 факторами: препарат: IA / IO / контроль, сторона: слева / справа, время: до / после обучения. Односторонний дисперсионный анализ ANOVA использовался для сравнения значений визуализации тензора диффузии и различий в BOLD-ответе между группами. Данные представлены как среднее + / — SEM, если не указано иное.
Для анализа динамики скорости обучения на протяжении всего периода обучения мы применили регрессионную модель, зависящую от фактора дополнительного кислорода.Используя регрессию наименьших квадратов, мы оценили экспоненциальную модель с тремя параметрами: нормализующей константой, экспоненциальным параметром «скорости обучения», который измеряет скорость обучения группы обработки воздуха, и переменной взаимодействия, которая измеряет разницу между скоростью обучения две лечебные группы. Для оценки модели мы использовали стандартный подход, описанный в 61 (страницы 483–485), который использует метод Гаусса – Ньютона (страницы 472–475).
Пусть γ 0 , γ 1 , γ 2 обозначают три параметра, описанные выше, соответственно.Наша модель выглядит следующим образом:
$$ Y_ {i} = \ upgamma _ {0} \ exp \ left ({\ upgamma _ {1} X_ {iT} + \ upgamma _ {2} X_ {iT} * X_ {iO2}} \ right) $$
где \ (X_ {T} \) — время (в днях), \ (X_ {O2} \) — индикаторная переменная для группы лечения O 2 и \ (Y \) — процент CR. Для нашего анализа мы в первую очередь рассматривали статистическую значимость параметра члена взаимодействия (\ (\ upgamma _ {2} \)). Мы построили 95% доверительный интервал для оценки значимости на пятипроцентном уровне.
Для нашего анализа мы в первую очередь рассматривали статистическую значимость параметра члена взаимодействия (\ (\ upgamma _ {2} \)). Мы построили 95% доверительный интервал для оценки значимости на пятипроцентном уровне.
Мэтью Деррик, Мэриленд | NorthShore
Лечащий врач
Педиатрия
Контакты
847.570 2920
847.570 0231
Факс
mailto: [email protected]
Образование
- Бакалавриат: 1980-83 Колледж Робинсона, Кембриджский университет, Англия, бакалавр с отличием по генетике, Массачусетс Кантаб.
- Медицинская школа: 1984-89 Лондонский госпитальный медицинский колледж, Лондонский университет, Уайтчепел, Лондон, E1, Англия. MBBS (июнь 1989 г.)
- Педиатрический Резидентура: 1991–1994 Детская Мемориальная больница, Северо-Западный университет, Чикаго, Иллинойс 60614, США.
- Неонатология Стипендия: 1995-1998 CVRI, UCSF, Сан-Франциско, Калифорния 94043, США
Сертификация Совета
- Американский совет педиатрии No.053104 12 октября 1994 г. Повторная сертификация 28 сентября 2001 г.

- Совет по неонатальной и перинатальной медицине Американского совета педиатрии № 003460 15 ноября 1999 г. Повторная сертификация 12 декабря 2006 г.
Исследования и академические интересы
Мое фундаментальное научное исследование было направлено на облегчение травм головного мозга у недоношенных детей в надежде, что понимание путей травм может быть использовано в будущих методах лечения человеческих младенцев.Как академический неонатолог, я расширил свои исследовательские интересы, включив в них клинические исследования открытого артериального протока и респираторного дистресс-синдрома, как главный исследователь в Northshore по многоцентровым исследованиям. Я также являюсь главным исследователем в Northshore в сотрудничестве с Дженнфер Баца в нескольких исследованиях, посвященных консультированию семей во время беременности. Мой план состоит в том, чтобы объединить фундаментальные науки с клиническими исследованиями для предотвращения и лечения перинатальных травм головного мозга.Моя общая карьерная цель — способствовать улучшению результатов жизни младенцев и их семей.
Опыт работы
- июл 98-июн 99 Клинический инструктор , Департамент педиатрии, UCSF, Сан-Франциско, Калифорния, 94143, США.
- июль 99 — июль 09 Доцент кафедры педиатрии Северо-Западного университета, Эванстон, Иллинойс.
- июль 2009 г. Доцент кафедры клинической практики Чикагского университета. Чикаго Иллинойс
- Июль 1999 г. Комитет по переливанию крови
- июль 2001 Комитет по перинатальной заболеваемости и смертности
- , январь 2011 г.
 , Система здравоохранения Нортшорского университета, IUCAC
, Система здравоохранения Нортшорского университета, IUCAC
Профессиональное членство / Принадлежность / Деятельность
- Член Американской академии педиатрии
Научная работа
Публикации в рецензируемых журналах:
- О’Дональд П., Деррик М. , Майерус М., Вейр Дж. Популяционно-генетическая теория вспомогательного спаривания, полового отбора и естественного отбора двухточечной божьей коровки, Adalia Bipunctata. Наследственность, 1984 52 (1) 433-461
- Collins J, Derrick M , Hilder L, Kempley S. Связь материнской этнической принадлежности с массой тела при рождении ребенка в Восточном Лондоне, Англия. Этническая принадлежность и болезни, 1997 7 1-4.
- Hawgood S, Derrick M , Poulain F. Структура и функция поверхностно-активного белка B.Biochimica et Biophysica Acta 1408 (1998) 150-160.
- Tan S, Bose R, Derrick M . Гипоксия-ишемия в мозге плода кролика увеличивает продукцию активных форм азота: количественная оценка нитротирозина. (2001) Free Radical Biol Med 30 (9) 1045-1051
- Brasch F, Ochs M, Kahne T., Guttentag S, Schauer-Vukasinovic V, Derrick M , Johnen G, Kapp N, Muller KM, Richter J. Giller T, Hawgood S, Buhling F. Вовлечение напсина A в C- и N-концевой процессинг сурфактантного белка B в пневмоцитах II типа легкого человека.(2003) J Biol Chem 278 (49) 49006-14
- Derrick M , Back, S, Ji X, Fisher K, Bregman J, Jilling T, Tan S. Преждевременная гипоксия-ишемия плода вызывает гипертонию и двигательные нарушения у новорожденных кроликов: модель церебрального паралича человека? (2004) Журнал неврологии 24 (1) 24-34
- Drobyshevsky A, Song S, Gamkrelidz G, Wyrwicz AM, Derrick M , Meng F, Li L, Ji X, Trommer, B, Beardsley DJ, Luo NL, Back SA, Tan S.
 Изменения в развитии диффузной анизотропии совпадают с прогрессирование незрелых олигодендроцитов и созревание потенциала действия соединения.(2005) J. Neurosci. 25 (25) 5988-97
Изменения в развитии диффузной анизотропии совпадают с прогрессирование незрелых олигодендроцитов и созревание потенциала действия соединения.(2005) J. Neurosci. 25 (25) 5988-97 - Tan S, MD; Дробышевский А., Джиллинг Т., Джи Х, Ульман Л.М., Энглоф I, Деррик М. Модель церебрального паралича у перинатального кролика. (2005) J Child Neurol 20 (12) 972-9 Derrick, M .; Дробышевский, А .; Ji, X .; Тан, С. Модель церебрального паралича в результате гипоксии-ишемии плода (2007 г.) Инсульт 38 (2) 731-5
- Дробышевский, А .; Derrick, M .; Wyrwicz, A.M .; Ji, X .; Englof, I .; Ullman, L.M .; Zelaya, M.E .; Нортингтон, Ф.J .; Tan, S. Повреждение белого вещества коррелирует с гипертонией на животной модели церебрального паралича. (2007) J Cereb Blood Flow Metab 27 (2) 270-81 .
- Дробышевский А., Деррик М., Прасад П.В., Джи Х, Энглоф И., Тан С. МРТ-ответ мозга плода, вызванный острой гипоксией-ишемией, прогнозирует постнатальный исход. Анналы неврологии, 2007, 61 (4): 307-314.
- Дробышевский А., Брегман Дж., Стори П., Мейер Дж., Прасад П.В., Деррик М., Маккендрик В., Тан С. Тензорная визуализация последовательной диффузии выявляет изменения белого вещества, которые коррелируют с двигательными исходами у недоношенных детей.Dev Neurosci, 2007; 29: 289–301.
- Ji, H .; Tan, S .; Igarashi, J .; Li, H .; Derrick, M .; Martasek, P .; Роман, Л. Дж .; Vasquez-Vivar, J .; Poulos, T. L .; Сильверман, Р. Б. Селективные нейрональные ингибиторы синтазы оксида азота и профилактика церебрального паралича. Анналы неврологии 2009 65 (2): 209-17
- Vásquez-Vivar, J., Whitsett, J., Derrick, M ., Ji, X., Yu, L., Tan, S. Тетрагидробиоптерин в профилактике гипертонии при гипоксии мозга плода Анналы неврологии 66 (3 ) 323-31 2009
- Деррик, M .; Дробышевский, А .; Ji, X .; Chen, L .; Ян, Й .
 ; Ji, H .; Silverman, R. B .; Тан, С. Гипоксия-ишемия вызывает стойкие двигательные нарушения в перинатальной модели церебрального паралича на кроликах: по оценке с помощью нового теста плавания. Int J Dev Neurosci 27 (6) 549-57 2009
; Ji, H .; Silverman, R. B .; Тан, С. Гипоксия-ишемия вызывает стойкие двигательные нарушения в перинатальной модели церебрального паралича на кроликах: по оценке с помощью нового теста плавания. Int J Dev Neurosci 27 (6) 549-57 2009 - Бузер, Дж. Р., Сеговия К. Н., Дин, Дж. М. Нельсон, К. Бердсли, Д. Гонг, X. Луо, Н. Л. Рен, Дж. Ван, И. Риддл, А. МакКлюр, М. М. Джи, X, Деррик, M ., Hohimer, AR., Back SA и S. Tan. Время появления предшественников поздних олигодендроцитов совпадает с повышенной восприимчивостью белого вещества головного мозга недоношенных кроликов к гипоксии-ишемии.J Cereb Blood Flow Metab. 30 (5) 1053-65
- Рао С. Лин З, Дробышевский А., Чен Л., Джи Х, Джи Х, Ян Й, Ю. Л., Деррик М., Сильверман Р. Б., Тан С. Вовлечение нейрональной синтазы оксида азота в продолжающуюся травму головного мозга плода после близкого к ней заболевания. Термин Гипоксия-Ишемия Кролика. Dev Neurosci, 2011. 33 (3-4): с. 288-98.
- Ю., Л. Деррик, М . Джи, Х. Сильверман, Р. Б. Уитсетт, Дж. Васкес-Вивар, Дж. Тан, С. Ингибирование нейрональной оксид-синтазы предотвращает церебральный паралич после гипоксии-ишемии у плодных кроликов: сравнение между JI-8 и 7-нитроиндазолом.Dev Neurosci, 2011. 33 (3-4): с. 312-9.
- Н. Дж. Робертсон, С. Тан, Ф. Грюнендал, Ф. ван Бел, С. Э. Юул, Л. Беннет, М. Деррик, С. А. Бэк, Р. К. Вальдес, Ф. Нортингтон, А. Дж. Ганн и К. Маллард. Какие нейропротекторные агенты готовы для трансляции от скамьи к постели у новорожденного? J Pediatr. 2012 8 февраля
- Л. Беннет, С. Тан, Л. Ван ден Хейдж, М. Деррик , Ф. Грюнендал, Ф. ван Бел, С. Юул, С. А. Бэк, Ф. Нортингтон, Н. Дж. Робертсон, К.Маллард и А. Дж. Ганн. Клеточная терапия неонатальной гипоксии-ишемии и церебрального паралича. Энн Нейрол. 2012 Май; 71 (5): 589-600.
- А. Дробышевский, К. Луо, М. Деррик, Л. Ю., Х. Ду, П. В. Прасад, Х. Васкес-Вивар, И.
 Батиник-Хаберле и С. Тан. Двигательный дефицит вызывается реперфузионно-реоксигенационным повреждением, диагностированным с помощью МРТ, и механизмом, включающим оксиданты. J Neurosci. 2012 18 апреля; 32 (16): 5500-9.
Батиник-Хаберле и С. Тан. Двигательный дефицит вызывается реперфузионно-реоксигенационным повреждением, диагностированным с помощью МРТ, и механизмом, включающим оксиданты. J Neurosci. 2012 18 апреля; 32 (16): 5500-9. - М. Деррик , И. Энглоф, А. Дробышевский, К. Луо, Л. Ю и С.Тан. Внутриутробная гибель плода может быть далека от провоцирующего инсульта на животной модели гипоксии-ишемии. Pediatr Res. 2012 11 мая.
- А. Дробышевский, М. Деррик , К. Ло, Л. Чжан, Ю. Ву, Л. Ю., С. Тан, Ближайшая гипоксия-ишемия плода у кроликов: МРТ может предсказать аномалии мышечного тонуса и глубокую гипоксию плода. Травма головного мозга. Ход 43 (10): 2757-2763.
- А. Дробышевский, Л. Ю., Ю. Ян, С. Халид, К. Ло, Р. Цзян, Х. Цзи, М. Деррик, , Л. Кей, Р. Б. Сильверман, С.Тан. Антенатальные инсульты изменяют обонятельную функцию новорожденных за счет оксида азота, продуцируемого нейрональной синтазой оксида азота. Exp Neurol 237 (2): 427-434.
- Р. Климан, А. Викремасингхе, Н. Джавери, Д. К. Хассинджер, Дж. Т. Аттридж, У. Санока, Р. Полин, М. Гиллам-Кракауэр, Дж. Риз, М. Маммел, Р. Кузер, Н. Малруни, Т. Д. Yanowitz, M. Derrick , P. Jegatheesan, M. Walsh, A. Fujii, N. Porta, WA Carey и JR Swanson. Энтеральное питание во время лечения незащищенного артериального протока индометацином и ибупрофеном.J Pediatr. 2013 5 марта Дата публикации: 12.03.2013.
- L. Yu, J. Vasquez-Vivar, R. Jiang, K. Luo, M. Derrick , S. Tan. Восприимчивость нейронов к преходящей тетрагидробиоптериновой недостаточности и антенатальной гипоксии-ишемии у плодов кроликов. Free Radic Biol Med 2014 67 426-36 Дата публикации: 2013/12/10.
- А. Дробышевский, Р. Цзян, Л. Линь, М. Деррик, , К. Ло, С. А. Бэк и С. Тан. : Потеря немиелинизированных аксонов с постнатальной гипертонией после гипоксии плода.Ann Neurol 2014 6 февраля.
Дата публикации : 19.03.2014
Главы книги:
- Реактивные формы митрогена и кислорода при перинатальной гипоксии-ишемии. Мэтью Деррик и Сидхартха Тан. Механизмы гипоксического повреждения головного мозга у новорожденных и потенциальные стратегии нейропротекции 2007.
Рецензент журнала для:
- Исследование мозга
- Нейробиология развития
- Свободная радикальная биология и медицина
- Журнал неврологии
- Журнал нейрохимии
- Нейрохимические исследования
- Педиатрические исследования
- Журнал нейротравм
Тезисы:
- Derrick M, Brinien L, Hawgood S.Гетерологичная экспрессия и сворачивание in vitro рекурсора человеческого поверхностно-активного белка B. Детское отделение 43 (4) A1642 1998
- Solano JP., Derrick, M., Sola A.: Вдыхаемый фрусомид не улучшает показатели функции легких у младенцев с хроническим заболеванием легких на ИВЛ. Журнал исследовательской медицины 1998 46: 1 (Abs)
- Бозе Р., Джиллинг Т., Каплан М., Деррик М., Тан С. Электрохимическое определение нитротирозина в мозге плода после повторяющейся ишемии матки. Free Radic Biol Med 27 (приложение 1): S73, 1999
- Brasch F, Ochs M, Derrick M , Johnen G, Wahlers T., Muller KM, Hawgood S, Richter J. Иммунолокализация ProSP-B и SP-B в легком человека. Амер. Jour Resp & Critical Care Medicine 161 (3) A44 2000
- Derrick M, Brady E, He J, Bose R, Kallio P, Tan S. Проточный цитометрический анализ in vitro судьбы эмбриональных клеток мозга кролика после острой гипоксии in vivo. Pediatr Res 47 (4): 396A, 2000.
- Derrick M, He J, Brady E, Bose R, Griffith OW, Babu RB, Tan S. In situ жизнеспособность эмбриональных клеток мозга кролика с ингибированием синтазы оксида азота после острой in vivo гипоксии-реоксигенации.
 Педиатр Res 47 (4): 457A, 2000.
Педиатр Res 47 (4): 457A, 2000. - Tan S, Jilling T, Derrick M , He J, Brady E, Bose R, Bregman J. Влияние антенатальной гипоксии-реоксигенации на нейроповеденческую оценку новорожденных кроликов. Педиатр Res 47 (4): 435A, 2000.
- Тан С., Бозе Р., Хе Дж., Брэди Е., Бабу Р., Гриффит О. В. .Derrick M, Увеличение количества реактивных видов оксида азота в мозге плода кроликов после гипоксии-реоксигенации. Педиатр Res 47 (4): 466A, 2000
- Tan S, Pouremad R, Wyrwicz A, He J, Brady E, Derrick M, Luo NL, Back SA. Повреждение белого вещества у плодов кроликов после внутриутробной гипоксии-ишемии приводит к гибели предшественников олигодендроцитов. Soc. Neurosci. Abstr., Vol 26, Part 2, p 2065, 2000.
- Tan S , Derrick M, He J, Brady E, Qu X-W, Wei H, Babu BR, Griffith OW.Ингибирование NOS нейронов снижает повреждение мозга плода кролика, опосредованное реактивными формами азота, после гипоксии . Free Radic Biol Med 29 Приложение 1: S77, 2000.
- Brasch F, Johnen G , Derrick M, Schauer-Vukasinovic V, Ochs M, Muller K, Richter J, Giller T, Hawggod S, Buhling F. Катепсин H и напсин A участвуют в переработке proSP-B. ATS 2001.
- Derrick M, Brady E, He J, Tan S. Кортикальная нейрональная травма плода кролика после острой повторяющейся реоксигенации маточной гипоксии.Педиатр Res 49 (4) 352A, 2001.
- Derrick M, He J, Brady E, Bregman J, Tan S. Фенотип церебрального паралича у новорожденных кроликов после дородовой гипоксии-ишемии у недоношенных плодов. Педиатр Res 49 (4) 432A, 2001.
- Tan S, Venkatasubramanian PN , Derrick M, Tom B, Wyrwicz AM, He J, Brady E, Luo NL, Back SA. In Utero Гипоксия-ишемия приводит к повреждению белого вещества и смерти предшественников олигодендроцитов у плодов кроликов. Педиатр Res 49 (4) 446A, 2001.

- TanS. Derrick M, Jilling T, Brady E, Chen J, Parks DA. Повреждение нейронов плода у недоношенных кроликов после ингибирования ксантина и повторяющейся гипоксии-ишемии. Soc. Neurosci. Abstr., Vol. 27, Программа № 572.14, 2001.
- Derrick M, Ji X, Fisher KM, Chen J, Crow JP, Tan S. In vitro спасение фетальных клеток мозга кролика антиоксидантами после in vivo гипоксически-ишемических инсультов. Педиатр Res 51 (4) 364A, 2002.
- Huynh N, Liu X, Goss D, Derrick M, Tan S.Преждевременная гипоксия-ишемия плодов кроликов увеличивает экспрессию нейрональной синтазы оксида азота. Педиатр Res 51 (4) 446A, 2002.
- Derrick M, Ji X, Huynh N, Fisher KM , Chen J, Tan S. In vitro оценка митохондриальной функции эмбриональных клеток мозга кролика после in vivo гипоксически-ишемических инсультов. Педиатр Res 51 (4) 446A, 2002.
- Derrick M, Ji X, Huynh N, Fisher KM , Chen J, Tan S. . Внутриутробный рост мозга и тела плода после единичного эпизода плацентарной недостаточности у недоношенного кролика.Педиатр Res 51 (4) 465A, 2002.
- M. Derrick, X. Ji, K. Fisher, J.- Bregman, T. Jilling, S. Tan. Антенатальная гипоксия-ишемия у недоношенных плодов приводит к фенотипу церебрального паралича и перинатальной смерти новорожденных кроликов. Программа № 488.4. 2002 Наблюдатель тезисов / Планировщик маршрута. Вашингтон, округ Колумбия: Общество неврологии, 2002.
- С. Тан, Н.- Хюин, К. Фишер, Х. Джи, Дж. П. Кроу, М. Деррик, . Роль нейрональной синтазы оксида азота и активных форм азота в повреждении головного мозга после гипоксии-ишемии плода кролика.Программа № 488.5. 2002 Наблюдатель тезисов / Планировщик маршрута. Вашингтон, округ Колумбия: Общество неврологии, 2002. Интернет
- Fisher K, Derrick M, Ji X, Shah N, Silverman RB, Hah JM, Tan S.
In vitro судьба клеток мозга плода с обработкой Rick1, нового ингибитора нейрональной синтазы оксида азота, после in vivo гипоксия-ишемия. Общество педиатрических исследований 2003 г., линия
- Derrick M, Fisher K, Ji X, Shah N, Tan S.Влияние фактора активации тромбоцитов на судьбу первичных диссоциированных клеток головного мозга плода. Общество педиатрических исследований 2003 г. в сети.
- Derrick M, Ji X, Fisher K, Tan S. Роль активных форм кислорода в продолжающемся повреждении клеток головного мозга эмбриональных кроликов после in vivo гипоксии / ишемии. Номер программы: 530.9 2003 Наблюдатель тезисов / Планировщик маршрута. Вашингтон, округ Колумбия: Общество неврологии, 2003. Интернет
- Тан С., Деррик М., Луо Н.Л., Брегман Дж. К., Джиллинг Т., Джи Х, Фишер К., Гладсон К.Л., Мердок Дж., Бэк С.Активированная реакция микроглии и гипертония при рождении на модели церебрального паралича, включающей преждевременную гипоксию-ишемию. Номер программы: 40.9 2003 Наблюдатель тезисов / Планировщик маршрута. Вашингтон, округ Колумбия: Общество неврологии, 2003. Интернет
- Derrick M , Ji X, Ullman LM, Kotlajich MV, Shah N, Tan S. Фактор активации тромбоцитов ацетилгидролаза (LIS-1) и развивающийся мозг изменяются с гипоксией-ишемией. Общество педиатрических исследований 2004 г., линия
- Дробышевский А., Бэк С.А., Вирвич А.М., Деррик М , Ли Л., Джи Х, Котлайич М.В., Тан С.Диффузионная тензорная визуализация развивающегося мозга кролика. Общество педиатрических исследований 2004 г. в сети.
- Derrick M , Ji X, Ullman L.M, Kotlajich MV, Tan S. Фактор активации тромбоцитов (PAF) и PAF-ацетилгидролаза (LIS-1) в развивающемся мозге кролика. Наблюдатель тезисов / Планировщик маршрута. Вашингтон, округ Колумбия: Общество неврологии, 2004. Интернет
- Дробышевский А., Бак С.А., Вирвич А.М., Деррик М. , Ли Л., Джи Х, Котлайич М.В., Тан С. Развитие белого вещества в перинатальном мозге кролика с помощью МРТ и электрофизиологии. Наблюдатель тезисов / Планировщик маршрута. Вашингтон, округ Колумбия: Общество неврологии, 2004. Интернет
- Drobyshevsky A, Derrick M , Ullman LM, Englof I, Ji X, Wyrwicz AM, Tan S. Травма белого вещества коррелирует с гипертонией в модели церебрального паралича. Общество педиатрических исследований 2005 г., линия
- Тан С., Дробышевский А., Джиллинг Т., Джи Икс, Ульман Л.М., Энглоф I, Деррик М . Гипертония сохраняется в модели церебрального паралича у перинатального кролика. Общество педиатрических исследований 2005 г., линия
- М.Derrick , A. Drobyshevsky, I. Englof, X. Ji, L.M. Ullman, S. Tan. Модифицированный тест принудительного плавания для количественной оценки стойких дефицитов в перинатальной модели церебрального паралича на кроликах. Программа № 603.17. 2005 Наблюдатель тезисов / Планировщик маршрута. Вашингтон, округ Колумбия: Общество неврологии, 2005. Интернет
- А. Дробышевский , М. Деррик , А. Wyrwicz, X. Ji, L. Ullman, I. Englof, S. Tan. Травма белого вещества коррелирует с гипертонией в модели церебрального паралича. Программа № 222.12. 2005 Наблюдатель тезисов / Планировщик маршрута.Вашингтон, округ Колумбия: Общество неврологии, 2005. Интернет
- Дробышевский А, Деррик М., АМ. Wyrwicz, X. Ji, I. Englof, ME. Селайя, Ф.Дж. Нортингтон, С. Тан Прогностическое значение показателей МРТ травмы белого вещества для гипертонии в модели церебрального паралича. Общество педиатрических исследований, 2006 г., линия
- А. Дробышевский, AM. Робинсон, М.Деррик, AM. Wyrwicz, X. Ji, I. Englof, S. Tan. Новое применение MEMRI выявляет сенсорный дефицит в результате повреждения обонятельной системы у новорожденного кролика после дородовой гипоксии-ишемии.Общество педиатрических исследований, 2006 г., линия
- M. Derrick , A.Drobyshevsky, X.Ji, I. Englof, S. Tan. Легкая двигательная дисфункция и тяжелые нарушения сохраняются, как показывает количественная оценка движений суставов в перинатальной кроличьей модели церебрального паралича. Общество педиатрических исследований, 2006 г., линия .
- Suma Rao, Zhenlang Lin, Xinhai Ji, Nina Sahay, Alexander Drobyshevsky, Lei Yu, Richard Silverman, Matthew Derrick , Sidhartha Tan. Ингибирование нейрональной синтазы оксида азота в срезах мозга плода кролика после травмы in vivo является защитным действием.Общество педиатрических исследований 2007 г., линия
- Мэтью Деррик , Ричард Б. Сильверман, Хайтао Цзи, Нина Сахай, Чжэнлан Линь, Синьхай Цзи, Сидхартха Тан. Введение нового ингибитора nNOS, JI-8 предотвращает проявление фенотипа церебрального паралича у плодов кроликов. Общество педиатрических исследований 2007 г., линия
- Александр Дробышевский, Мэтью Деррик , Алиса М. Вырвиц, П.В. Прасад, Синьхай Джи, Нина Сахай, Сидхартха Тан. Диффузионно-взвешенная визуализация предсказывает фенотип церебрального паралича после гипоксии-ишемии плода у кроликов.Общество педиатрических исследований 2007 г., линия
- Чжэнланг Линь, Эмили Обринджер, Нина Сахай, Синьхай Цзи, Сидхартха Тан, Мэтью Деррик. Плоды самцов и самок кроликов не показали различий в повреждении клеток головного мозга после гипоксии-ишемии плода. Общество педиатрических исследований, 2007 г., линия .
- A. Дробышевский, J. Vasquez-Vivar, L. Yu, N. Sahay, Z. Lin, X. Ji, M. Derrick , S. Tan. Может ли оксидативный стресс объяснить острые изменения на МРТ, которые напрямую связаны с возможным двигательным фенотипом церебрального паралича.Общество нейробиологии, 2007. Интернет
- М. Деррик , Дж. Уитсетт, Л. Ю, Х. Джи, С. Тан, Дж. Васкес-Вивар. Добавление тетрагидробиоптерина в модель церебрального паралича в Обществе педиатрических исследований кроликов, 2008 г., линия
- Лей Ю, Синьхай Цзи, Александр Дробышевский, Мэтью Деррик, Тамас Джиллинг, Сидхарта Тан. Молекулярное клонирование кроличьей ксантиноксидазы и развитие модуляции ее экспрессии в мозге плода. Общество педиатрических исследований, 2008 г., линия
- Керст Дж.Нельсон, Кристен Н. Сеговия, Мелисса МакКлюр, Мэтью Деррик, , Сидхарта Тан, Стивен А. Бэк. Масштабы повреждения белого мозга плода кролика от гипоксии-ишемии совпадают со сроками появления предшественников поздних олигодендроцитов (OL). Общество педиатрических исследований, 2008 г., линия
- Александр Дробышевский, Мэтью Деррик , Синьхай Цзи, Лей Ю, Сидхартха Тан. Новое применение МРТ с усилением марганца показывает, что базальные ганглии ведут себя по-разному у новорожденных кроликов с гипертонией после дородовой гипоксии-ишемии.Общество педиатрических исследований 2008 г. в сети.
- Лей Ю, Александр Дробышевский, Синьхай Цзи, Мэтью Деррик , Сидхарта Тан. [506] Уровень нейрональной синтазы оксида азота является критическим детерминантом исследования Педиатрического общества по выживанию нейронов, 2008 г. онлайн.
- Жанетт Васкес-Вивар, Дженнифер Уитсетт, Синьхай Цзи, Лей Ю, Мэтью Деррик, , Сидхарта Тан. Нейропротекторное действие тетрагидроптерина в отношении двигательных нарушений в мозге плода, вызванных гипоксией.Шестая конференция Херши по развивающимся травмам головного мозга.
- Лэй Ю, Александр Дробышевский, Синьхай Цзи, Мэтью Деррик . Уровень нейрональной синтазы оксида азота является критическим фактором гибели нейронов после гипоксии-ишемии плода. Шестая конференция Херши по развивающимся травмам головного мозга.
- Лей Ю, Синьхай Цзи, Александр Дробышевский, Мэтью Деррик, , Тамас Джиллинг, Сидхартха Тан. Молекулярное клонирование кроличьей ксантиноксидазы и изменение ее экспрессии в мозге плода в процессе развития.Шестая конференция Херши по развивающимся травмам головного мозга.
- Л.Ю., X.JI, А.Дробышевский, М. ДЕРРИК , С.ТАН. Молекулярное клонирование кроличьей ксантиндегидрогеназы проясняет паттерн развития экспрессии в перинатальном мозге и ответ на гипоксию-ишемию. Общество неврологии, 2008. Интернет
- M. DERRICK , L. YU, A. DROBYSHEVSKY, X. JI, S. TAN Уровень нейрональной синтазы оксида азота является критическим фактором гибели нейронов после гипоксии-ишемии плода. Society for Neuroscience, 2008 Online
- Синьхай Цзи, Элизабет Трейси, Александр Дробышевский, Мэтью Деррик , Лэй Ю, Айпин Лю, Майкл Коттен, Рональд Н.Гольдберг, Джоан Керцберг, Сидхарта Тан. Улучшают ли клетки пуповинной крови человека исход в модели церебрального паралича у плода кролика. 2009 Наблюдатель тезисов / Планировщик маршрута. PAS, встреча SPR в Балтиморе, Мэриленд. 3 мая 2009 г. Стендовый симпозиум
- Лей Ю, Синьхай Цзи, Мэтью Деррик , Сидхарта Тан, Александр Дробышевский. Ответ обонятельной системы новорожденного кролика in vivo на стимул оксида азота. 2009 Наблюдатель тезисов / Планировщик маршрута. PAS, встреча SPR в Балтиморе, Мэриленд.3 мая 2009 года. Плакат
- Александр Дробышевский, Лайсян Линь, Мэтью Деррик , Синьхай Цзи, Лэй Ю, Сидхартха Тан . Структурно-функциональные отношения путей белого вещества по данным МРТ и электрофизиологии в модели церебрального паралича у кроликов. 2009 Наблюдатель тезисов / Планировщик маршрута. PAS, встреча SPR в Балтиморе, Мэриленд. 3 мая 2009 года. Плакат
- Мэтью Деррик , Александр Дробышевский, Синьхай Цзи, Лина Чен, Йиро Ян, Сидхартха Тан. Анализ движения суставов во время поддерживаемого плавания выявляет различия у кроликов с негипертонической гипоксией и ишемией и изменения с помощью очень селективного ингибитора nNOS.2009 Наблюдатель тезисов / Планировщик маршрута. PAS, встреча SPR в Балтиморе, Мэриленд. 3 мая 2009 года. Плакат
- L. Yu 1 , A. Drobyshevsky, X. Ji, M. Derrick , S. Tan. УРОВЕНЬ СИНТЕЗА НЕЙРОНА ОКСИДА АЗОТА ЯВЛЯЕТСЯ КРИТИЧЕСКИМ ДЕТЕРМИНАНТОМ НЕЙРОННОЙ СМЕРТИ ПОСЛЕ ГИПОКСИИ-ИШЕМИИ плода . XXIV Международный симпозиум по церебральному кровотоку и метаболизму и IX Международная конференция по количественной оценке функции мозга с помощью ПЭТ Чикаго, Иллинойс, США — 29 июня — 3 июля 2009 г.
- А.Дробышевский, М. Деррик , X. Ji, L. Yu, S. Tan. РЕГИОНАЛЬНАЯ ХАРАКТЕРИСТИКА И ВРЕМЕННОЕ ТЕЧЕНИЕ ТРАВМЫ ПРИ БЛИЗНЕЙШЕЙ ГИПОКСИИ-ИШЕМИИ ПЛОДА КРОЛИКА. XXIV Международный симпозиум по церебральному кровотоку и метаболизму и IX Международная конференция по количественной оценке функции мозга с помощью ПЭТ Чикаго, Иллинойс, США — 29 июня — 3 июля 2009 г.
- М. Деррик , А. Дробышевский, X. Ji, Y. Yang, R. Silverman, H. Ji, S. Tan. НОВЫЙ ПЛАВАТЕЛЬНЫЙ ТЕСТ ОПРЕДЕЛЯЕТ ДЕФИЦИТ ДВИЖЕНИЯ У ПЕРИНАТАЛЬНЫХ КРОЛИКОВ ПОСЛЕ ГИПОКСИИ-ИШЕМИИ ПЛОДА.XXIV Международный симпозиум по церебральному кровотоку и метаболизму и IX Международная конференция по количественной оценке функции мозга с помощью ПЭТ Чикаго, Иллинойс, США — 29 июня — 3 июля 2009 г.
- X. Ji ,, M. Derrick , L. Yu, A. Drobyshevsky, A. Liu, S. Tan. ФЕНОТИП ЦЕРЕБРАЛЬНОГО ПАРАЛИКА У КРОЛИКОВ, МОДЕЛИРОВАНИЕ ПРОГРЕССИИ ОТ ЧАСТИЧНОЙ ДО ПОЛНОЙ ПЛАЦЕНТАРНОЙ НЕДОСТАТОЧНОСТИ. XXIV Международный симпозиум по церебральному кровотоку и метаболизму и IX Международная конференция по количественной оценке функции мозга с помощью ПЭТ Чикаго, Иллинойс, США — 29 июня — 3 июля 2009 г.
- Синьхай Цзи, Алексендра Дробышевский, Хайтао Цзи, Мэтью Деррик , Лэй Ю, Айпин Лю, Ричард Б.Сильверман, Сидхартха Тан . Терапевтическое применение нового селективного нейронального ингибитора синтазы оксида азота при усовершенствованном моделировании фенотипа кролика с церебральным параличом от частичной до полной плацентарной недостаточности. 2010 Abstract Viewer / Планировщик маршрута. PAS, встреча SPR в Ванкувере, Калифорния. Май 2010 г.
- Лей Ю, Синьхай Цзи, Айпин Лю, Александр Дробышевский, Ронна Дорнсиф, Мэтью Деррик, Йиронг Ян, Майкл Коттен, Рональд Голдберг, Джоанн Курцберг и Сидхарта Тан.Определение безопасной и эффективной дозы клеток пуповинной крови человека в модели церебрального паралича у плода кролика. Седьмая конференция Херши по проблемам развития черепно-мозговой травмы Snowbird, 3-5 июня 2010 г.,
- Лей Ю, Кейт Заппиа, Мэтью Деррик , Бенджамин Карни, Синьхай Цзи, Дженнифер Уитсетт, Сидхарта Тан. Жизненно важная роль тетрагидробиоптерина в выживании нейронов в различных областях мозга плода . Седьмая конференция Херши по вопросам развития травм головного мозга Snowbird, 3-5 июня 2010 г.
- Александр Дробышевский, Синьхай Цзи, Иронг Ян, Мэтью Деррик, , Лей Ю, Сидхартха Тан.Снижение функциональной связи в наборах для послеродовых кроликов с двигательными нарушениями после дородовой гипоксии-ишемии . Седьмая конференция Херши по вопросам развития травм головного мозга Snowbird, 3-5 июня 2010 г.
- М. Деррик , Дж. Кондапалли, П.Т. Шумакер. Гипотермия и снижение гипоксии во время реперфузии. Уменьшение гибели клеток во время симулированной ишемии-реперфузии, аннотация № 754399, 2011 г., PAS, 30 апреля — 3 мая 2011 г., Денвер, Колорадо.
- М. Деррик , Джоанн Брегман, W.Маккендрик. Отличаются ли младенцы 23 недель от детей 24 и 25 недель ?, аннотация № 754295 PAS 2011, 30 апреля — 3 мая 2011 г., Денвер, Колорадо,
- L. Yu, A. Drobyshevsky, K. Luo, R. Jiang, M. Derrick , S.Tan. Реперфузионная травма Снижение количества олигодендроцитов в мозге плода кролика после гипоксии ишемии матки [1526.450] PAS Boston 2012
- Л.Ю., М. Деррик, К. Ло, Р. Цзян, А. Дробышевский, С. Тан. Бесполезность дизайна и методов лечения критического окислительного стресса для предотвращения двигательного дефицита в модели на животных [4501.26] PAS Boston 2012
- К. Луо, Л.Ю, А.Шах, Р.Цзян, А.Дробышевский , М. Деррик, С.Тань. Новая модель хориоамнионита у кроликов вызывает нейроповеденческий дефицит [1540.574] PAS 2013 Вашингтон, округ Колумбия, май 4 2013
- Дж. Батца, А. Жанвье, М. Деррик. Он сказал, что сказала: Предполагаемые и фактические опасения потенциальных родителей с угрозой преждевременных родов при минимальной жизнеспособности [2909.88] PAS 2013 Вашингтон, округ Колумбия 5 мая 2013 г.
- А.Дробышевский, Р. Цзян, Л. Линь, Л. Ю, М. Деррик , К. Ло, С.А. Бэк, С. Тан . Послеродовая гипертония, вызванная повреждением белого вещества в немиелинизированных трактах волокна после дородовой гипоксии-ишемии. PAS 2013 Вашингтон, округ Колумбия 7 мая 2013 г.
Гранты:
- PI. Стипендия по научно-исследовательской подготовке 7 / 97-6 / 98
- Американская ассоциация легких в Калифорнии. 20 000 долл. США
- Стипендия по исследовательской подготовке PI 7 / 98-6 / 99 Американская ассоциация легких.$ 35 000
- Главный исследователь. Программа пилотных грантов 6 / 04-5 / 05
- Исследовательский институт ENH. Роль фактора активации тромбоцитов и ацетилгидролазы PAF в развитии мозга. $ 29 750
- Co- Inv R-21 NS41476 Задняя часть (PI), вышка (Co-I) 8/1 / 01-7 / 31/04
- NINDS Клеточные механизмы преждевременной травмы белого вещества
- Co-Inv R01-NS043285-01A1 Tan (PI), Derrick (Co-I) 1.12.2002 — 30.11.2007
- NINDS / NICHD Реактивные виды азота при гипоксической травме головного мозга
- Co-Inv R01 NS051402 Тан (PI), вышка (Co-I) 07/1 / 06-06 / 30/11
- NINDS Развитие белого вещества в перинатальном периоде и гипоксия плода
- Co-Inv R21 R21HD054742-01 Тан (PI) 09/20 / 07-09 / 19/09
- NICHD Редокс-модуляция оксида азота при обонятельной дисфункции после гипоксии плода
- Co-Inv 2R56NS043285-06A2 Tan (PI) 8/09 / 10-09 / 08/11
- NINDS Нейрональная синтаза оксида азота при раннем критическом повреждении головного мозга плода в результате гипоксии
- PI K08- HD057307-01A1 1 сентября 2008 г. 31 августа 2013 г.
- NICHD Митохондрии, супероксид и MnSOD в модели гипоксического церебрального паралича плода
Публикации
Микробиота влияет на развитие мозга и поведение мышей C57BL / 6J.
Образец цитирования: Lu J, Synowiec S, Lu L, Yu Y, Bretherick T, Takada S, et al.(2018) Микробиота влияет на развитие мозга и поведение мышей C57BL / 6J. PLoS ONE 13 (8): e0201829. https://doi.org/10.1371/journal.pone.0201829
Редактор: Эфтимиос М. К. Скулакис, Исследовательский центр биомедицинских наук Александр Флеминг, ГРЕЦИЯ
Поступила: 30 апреля 2018 г .; Одобрена: 23 июля 2018 г .; Опубликовано: 3 августа 2018 г.
Авторские права: © 2018 Lu et al.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Все соответствующие данные находятся в документе и его файлах с вспомогательной информацией.
Финансирование: Эта работа была поддержана NIH 1R01HD083481 (EC). Спонсор не принимал участия в планировании исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявляют, что у них нет конкурирующих интересов.
Введение
Микробные сообщества в кишечнике младенца или кишечная микробиота все чаще рассматриваются как модифицируемый фактор, влияющий на развитие мозга и поведения хозяина [1–4]. Микробиота приобретается около рождения и развивается до относительно стабильного сообщества одновременно с развитием нервной системы в течение первых 2–3 лет и может оказывать прямое и глубокое влияние на познание и поведение в более позднем возрасте [5].Было показано, что кишечная микробиота участвует в раннем программировании мозговых цепей, которые опосредуют реакцию на стресс, двигательную активность, тревожное поведение и когнитивные функции в раннем детстве, а также потенциальный патогенез нарушений нервного развития, таких как аутизм, дефицит внимания. / расстройство гиперактивности и шизофрения [6–11].
В течение первых двух лет постнатальной жизни в нервной системе происходят глубокие изменения: массивное разрастание дендритов и аксонов, синаптогенез, разрастание глиальных клеток и миелинизация [12, 13].Объемные исследования на основе количественной магнитно-резонансной томографии (МРТ) неизменно показывают, что морфология мозга динамически изменяется от рождения до подросткового возраста и во взрослом возрасте [14, 15]. Исследования продемонстрировали общую положительную корреляцию между объемом серого вещества и показателями IQ у здоровых детей и подростков от 5 до 17 лет [16], а также у взрослых [17]. Кроме того, возрастное расширение и миелинизация белого вещества являются важными показателями созревания мозга и когнитивных функций в более позднем возрасте [18].
Польза комменсальной микробиоты на физиологию хозяина и на функцию и развитие мозга изучается с использованием мышей без микробов (GF) [19], демонстрируя важность микробиоты в регулировании множественных процессов развития мозга, включая синаптогенез и связанные с ним пути вторичных мессенджеров [6 , 20], миелинизация [21], серотонинергическая система гиппокампа [7], проницаемость гематоэнцефалического барьера [22], а также реакция оси гипоталамус-гипофиз-надпочечники на стресс [9], а также исследовательское и тревожное поведение [ 23].Исследования с использованием стереологической объемной оценки выявили региональные объемные изменения, такие как увеличение миндалевидного тела и гиппокампа, приписываемые уменьшению ветвления дендритов у взрослых мышей, свободных от микробов [24]. Однако связь между структурой мозга и профилями микробиоты не была описана у мышей с использованием методов нейровизуализации МРТ. Применение неинвазивных и высокопроизводительных методов МРТ было бы очень полезно для изучения влияния микробиоты на развитие структуры мозга, поскольку изменения часто незначительны, распределяются по мозгу и развиваются со временем.Кроме того, влияние микробиоты на мозг, вероятно, затронет несколько областей и клеточных мишеней, но большинство текущих исследований сосредоточено на одной или двух конкретных областях и типах поведения. В этом исследовании мы специально использовали объективные количественные методы нейровизуализации для всего мозга, чтобы оценить микроструктурные и объемные различия между мышами с комменсальными бактериями, также известными как мыши, свободные от специфических патогенов (SPF) и GF, в возрасте четырех недель (считающихся ювенильными) и Возраст 12 недель (считается взрослым).
В свете интригующих сообщений о нарушениях миелинизации в префронтальной коре мышей GF [21] мы уделили особое внимание оценке миелинизации и развития белого вещества в юношеском и взрослом возрасте. Миелинизация обеспечивает эффективную передачу информации между нейронными областями постнатально и считается важным индикатором созревания мозга [18]. Новый метод количественной МРТ in vivo , картирование фракции макромолекулярных протонов (MPF), был использован для реконструкции параметрических карт относительного количества макромолекулярных протонов, вызывающих эффект переноса намагниченности, который обеспечивает биомаркер миелинизации в нервных тканях [25, 26] .Метод был проверен на гистологических маркерах миелина в нормальном мозге крысы [27] и на мышиной модели демиелинизации [28]. Этот метод имеет высокое разрешение, быстр и не зависит от направленной организации ткани, что дает уникальную возможность оценить миелинизацию в структурах серого вещества [28, 29] и предоставляет дополнительную информацию для визуализации тензора диффузии в белом веществе, которая также используется в этом исследовании. .
Понимание того, как изменения в микробиоте кишечника в раннем возрасте способствуют уязвимости к поведенческим и когнитивным расстройствам, имеет первостепенное значение в клинической и поведенческой нейробиологии.Целью этого исследования было изучить вклад кишечных бактерий в поведение и связанные с ним структуры и функции мозга с использованием штамма C57BL / 6J мышей SPF и GF. Поскольку ожидается влияние как половой, так и возрастной микробиоты на мозг [30, 31], была проведена систематическая характеристика морфологических и миелинизационных изменений по всему мозгу между самцами и самками у молодых и взрослых мышей (в возрасте четырех и 12 недель). . Наши данные демонстрируют, что комменсальные бактерии необходимы для нормального морфологического развития и созревания в сером и белом веществе областей мозга с последствиями для поведенческих результатов, таких как локомоция и когнитивные функции.
Материалы и методы
Животные и общий план опытов
Исследование получило одобрение Институционального комитета по уходу за животными и использованию комитетов системы здравоохранения Университета Северного берега (NorthShore) в соответствии с протоколом Eh26-264 и Чикагского университета в соответствии с протоколом № 71703, и все исследования проводились строго в соответствии с требованиями США. Политика Службы общественного здравоохранения в отношении гуманного ухода за лабораторными животными и их использования, а также утвержденные правила ухода за животными и их использования в Чикагском университете.Свободных от зародышей (GF) мышей C57BL / 6J, первоначально полученных из лаборатории Джексона (Бар-Харбор, штат Мэн), разводили и содержали в гнотобиотическом учреждении Центрального центра исследования пищеварительных заболеваний при Чикагском университете. Колония GF регулярно проверялась на микробы и паразиты персоналом учреждения, чтобы гарантировать отсутствие микробов. Мыши C57BL / 6J, свободные от специфических патогенов (SPF), были получены из лаборатории Джексона. Мышей тестировали в двух временных точках развития: четыре недели (15 самцов и 15 самок SPF, девять самцов и шесть самок GF) и 12 недель (13 самцов и 13 самок SPF, семь самцов и семь самок GF).Для каждой временной точки использовались отдельные когорты. Не было разницы в массе тела между мышами SPF и GF одного пола (таблица 1). Животных перевозили на Северный берег в стерильных контейнерах за 3-5 дней до тестирования, чтобы обеспечить период акклиматизации. По прибытии в NorthShore мышей помещали в индивидуально вентилируемые клетки с HEPA-фильтром до начала поведенческого тестирования. Водный лабиринт Морриса выполняли на 1–5 сутки. В дни 1–5 мы начали с водного лабиринта Морриса в 10 часов утра.Животным давали возможность восстановиться в течение двух часов перед тем, как во второй половине дня на 1, 2 и 3 день выполняли тест в открытом поле, приподнятом крестообразном лабиринте или на социальное взаимодействие. Кондиционирование страха проводили на 7 и 8 дни. После поведенческого тестирования мышей подвергали объемной и макромолекулярной протонной фракции in vivo магнитно-резонансной томографии (МРТ) на 9 и 10 дни. Сразу после in vivo МРТ мышей подвергали глубокой анестезии. с пентобарбиталом натрия (100 мг / кг) и транскардиально перфузировали 30 мл фосфатно-солевого буфера (PBS) (pH 7.4) при комнатной температуре (25 ° С). После этого была проведена инфузия 30 мл 4% параформальдегида (PFA) в PBS с 2 мМ ProHance (Bracco Diagnostics Inc., Princeton, NJ). После перфузии головы были удалены вместе с кожей, нижней челюстью, ушами и хрящевым кончиком носа. Оставшимся структурам черепа, содержащим мозг, позволяли постфиксировать в 4% PFA при 4 ° C в течение ночи. Образцы переносили и хранили в PBS и 2 мМ растворе ProHance и 0,01% азида натрия до тех пор, пока они не подверглись ex vivo объемной и диффузионно-взвешенной МРТ высокого разрешения.После визуализации мозг был обработан для иммуногистохимии.
Магнитно-резонансная томография (МРТ)
Получение изображений фракции протонов макромолекул (MPF) in vivo .Визуализацию выполняли на спектрометре Bruker Avance 14,1 Т (Bruker, Billerica, MA) с использованием 20-мм резонатора. Мышей успокаивали ингаляцией изофлураном (Abbot, IL), разбавленным воздухом до 5% для индукции и 1,5% для поддержания. Частоту дыхания животных и ректальную температуру контролировали с помощью физиологического монитора мелких животных (SAII’s Small Animal Instruments, Нью-Йорк).Температуру тела поддерживали на уровне 35 90 283 o 90 284 ° C, поддерживая температуру окружающей среды в стволе на уровне 32 90 283 o 90 284 ° C с использованием контроллера градиента температуры спектрометра. Карты 3D MPF (рис. 1A) были получены из трех исходных изображений (передача намагниченности (MT) -, плотность протонов (PD) — и T1-взвешенное) с использованием одноточечного метода с синтетическим эталонным изображением [32]. PD- и T1-взвешенные изображения GRE были получены с TR / TE = 16 / 2,6 мс и α = 3 ° и 16 °, соответственно. МТ-взвешенные изображения были получены с TR / TE = 25/2.6 мс и α = 9 °. Импульс внерезонансного насыщения подавался на частоте смещения 6 кГц с эффективным углом переворота насыщения 500 °. Все изображения были получены в аксиальной плоскости с охватом всего мозга и разрешением 0,125×0,125×0,25 мм 3 . Все изображения были получены с четырьмя средними значениями сигнала и общим временем сканирования 33 мин. Во всех экспериментах по построению трехмерных изображений использовались линейный порядок фазового кодирования со 100 фиктивными сканированиями, избирательное возбуждение слябов и фракционное (75%) получение k-пространства в направлении выбора сляба.Для коррекции неоднородностей поля 3D-карты B0 и B1 были получены с использованием двойного TE (TR / TE1 / TE2 = 20 / 2,9 / 5,8 мс, α = 8 °) и визуализации с фактическим углом поворота (AFI) (TR1 / TR2). / TE = 13/65/4 мс, методы α = 60 ° соответственно [33]. Все процедуры реконструкции были выполнены с использованием специально написанного программного обеспечения на языке C.
Рис. 1. Методы МРТ.
(А). Карта фракции макромолекулярных протонов четырехнедельной мыши. Цветная полоса указывает псевдоцветное отображение MPF в процентных единицах. (В). Т1-взвешенные объемы высокого разрешения изображений мозга мышей ex -vivo с черепами in situ.Одновременно были получены изображения группы из четырех голов мозга. (С). Автоматическое структурное разбиение мозга мыши, полученное из изображения МТ с использованием метода слияния мультиатласных меток.
https://doi.org/10.1371/journal.pone.0201829.g001
Ex vivo МРТ.Ex vivo МРТ-визуализация проводилась от семи до 21 дня после фиксации. Черепа погружали в неводную среду (fomblin Y, Sigma Aldrich, MO). Визуализацию проводили на спектрометре Bruker Avance 14,1 Тл (Bruker, Биллерика, Массачусетс) с использованием 30-мм резонатора.Одновременно были визуализированы четыре мозга (рис. 1B). Т1-взвешенное изображение высокого разрешения для морфометрии на основе тензор было получено с параметрами TR / TE / NEX 17 / 4.8 / 8 с изотропным разрешением 60 мкм. Время съемки составляло четыре часа десять минут.
Эксперименты по визуализации тензорной диффузии (DTI) состояли из 30 неколлинеарных направлений диффузионных взвешенных спин-эхо-изображений с b = 0 и 1,0 мс / мкм 2 , δ / Δ = 3/7 мс. Параметры изображения: TR / TE / NEX 7500 / 15,1 / 1, 40 аксиальных срезов толщиной 0,25 мм без зазора, покрывающего весь головной мозг.Разрешение в плоскости составляло 125 x 125 мкм, а объемы мозга были интерполированы до изотропных 125 мкм 3 . Время съемки составило 11 часов 22 минуты. Карты тензора диффузии были рассчитаны с использованием многомерной линейной аппроксимации ослабления сигнала из полученных диффузно-взвешенных изображений [34]. Карты фракционной анизотропии (FA) были рассчитаны [35] с использованием собственного программного обеспечения, написанного на Matlab (MathWorks, Natick, MA).
Обработка данных МРТ.
Изображения с переносом намагниченности мозга мыши, полученные как компонент эксперимента in vivo MPF и обладающие отличным контрастом белого / серого вещества, были использованы для автоматического структурного разделения с использованием метода слияния мультиатласных меток, как подробно описано в [36 ].Вкратце, отдельные изображения головы мыши были обработаны для извлечения мозга, коррекции неоднородности интенсивности, аффинной регистрации с изображениями атласа и слияния этикеток. Использовалась общедоступная база данных атласа MRMNeAt, содержащая 10 индивидуально помеченных C57BL / 6J головного мозга мышей in vivo . 21 структура белого и серого вещества из атласа была помечена (рис. 1C) с добавлением области интереса префронтальной коры, которая была определена как кора на расстоянии 2 мм от начала головного мозга. Объемы отдельных анатомических областей и значения MPF сравнивались между группами SPF и GF.
Тензорный морфометрический анализ был выполнен на ex vivo T1-взвешенных изображениях высокого разрешения с использованием программ FSL (http://www.fmrib.ox.ac.uk/fsl/). Шаблоны для конкретных исследований были созданы для каждого изучаемого возраста путем итеративной регистрации головного мозга с обнаженным черепом (как описано выше для in-vivo изображений) случайным образом выбранных пяти GF и пяти SPF головного мозга. Для каждого возраста мозг регистрировали по аффинной схеме, а случайно выбранный мозг соответствующей возрастной группы объединяли и усредняли.Затем это усредненное изображение было перевернуто по оси x, и два зеркальных изображения повторно усреднены для получения первого прохода, специфичного для исследования «аффинного» шаблона. Во-вторых, мозг был повторно зарегистрирован в этом «аффинном» шаблоне с использованием нелинейной регистрации, усреднен и перевернут по оси абсцисс. Затем оба зеркальных изображения были усреднены для создания окончательного симметричного, специфичного для исследования «нелинейного» шаблона. Весь мозг (пять самцов и пять самок мышей SPF и GF для четырех- и 12-недельных групп) нелинейным образом регистрировали в шаблоне исследования / возраста.Для каждого воксела был вычислен натуральный логарифм определителя Якоби (JD) поля деформации, и объемы были сглажены с помощью ядер Гаусса, сигма = 0,1 мм. JD — это мера деформации каждого вокселя относительно изображения атласа. Его можно представить как величину, на которую необходимо умножить громкость этого воксела, чтобы достичь консенсусного среднего. JD> 1 означает расширение, а <1 означает уменьшение объема этого вокселя по сравнению с объемом того же вокселя в изображении атласа.Непараметрический вывод на основе перестановок между группами SPF и GF был выполнен на JD с использованием процедуры рандомизации FSL и опций расширения кластера без порога для управления частотой ошибок в семье.
Для анализа данных ex vivo DTI был использован межобъектный анализ пространственной статистики на основе вокселей (TBSS) [37, 38], реализованный в программном обеспечении FSL (http: //www.fmrib. ox.ac.uk/fsl/). Все индивидуальные объемы FA были зарегистрированы в шаблоне, средняя карта FA была рассчитана и прорежена, чтобы представить средний скелет FA.Для каждого субъекта данные вокселей проецировались с карт FA на ближайшие воксели на среднем скелете FA. Значения вокселей скелета были присвоены максимуму проецируемых вокселей. Значения вокселов на общем скелете были проанализированы с помощью воксельного межсубъектного статистического анализа с использованием общей линейной модели [37, 38]. В случае двух групп, как в этом исследовании, проверка контраста между групповыми предикторами эквивалентна непарному t-критерию средней разницы между группами.В результате процедуры были созданы статистические параметрические карты, содержащие p -значений для воксельных двухвыборочных непарных t-критериев. Результаты были скорректированы для множественных сравнений, контролируя частоту ошибок в семье. Для подпрограммы FSL непараметрического вывода перестановок использовались безпороговое расширение кластера и 3000 перестановок.
Миелиновая окраска
Оценка содержания миелина была выполнена на одной из серий толщиной 20 микрон с использованием красителя Luxol Fast Blue (LFB) (Sigma-Aldrich, Сент-Луис, Миссури).Мозг извлекали из черепов, подвергали криозащите 30% сахарозой и замораживали на сухом льду. Последовательные срезы толщиной 20 микрон разрезали на 400 микрон на криостате и помещали на предметные стекла, покрытые поли-L-лизином (Sigma-Aldrich). Все слайды, состоящие из пяти мозгов на группу (мужчины и женщины, SPF и GF, всего 40 слайдов), обрабатывались как одна партия с идентичными настройками. Срезы инкубировали в 0,1% растворе LFB в течение двух часов при 60 ° C, дифференцировали путем погружения в 0,05% раствор карбоната лития на 20 секунд, с продолжением дифференциации путем многократного погружения в 70% спирт до появления контраста серого вещества.Срезы промывали, обезвоживали в спирте, очищали в ксилоле и монтировали. Каждую серию окрашенных LFB всего мозга фотографировали с помощью цифровой камеры с разрешением восемь мегапикселей с использованием идентичных параметров изображения. Изображения были проанализированы с помощью программного обеспечения ImageJ (National Institutes of Health, Bethesda, MD). Интенсивность изображений в шкале серого измерялась в нескольких областях серого и белого вещества стандартного размера и стандартного расположения: 1,7 брегма — для лобной коры, 0 брегма — для передней комиссуры, полосатое тело, -1.28 брегма для фимбрии, мозолистого тела, внутренней капсулы, гиппокампа, таламуса и теменной коры. Наконец, интенсивности были инвертированы и нормализованы к средней интенсивности всех секций мозга.
Поведенческие исследования
Тестирование проходило в специальной тихой комнате. Мышам давали возможность акклиматизироваться по крайней мере в течение трех дней после отправки. Камеры для испытаний очищали 70% этанолом и проветривали в течение 3 мин после каждого животного. Движения животных регистрировались и обрабатывались с помощью программного обеспечения ЛЮБОГО лабиринта (Stoelting Co., Вуд Дейл, Иллинойс).
Испытание в открытом поле.
Животных помещали по отдельности в центр открытого ящика с чистым полем (61 × 61 см) и регистрировали их спонтанную двигательную активность. Компьютерная программа автоматически записывала следующие параметры: время в пути, средняя скорость, пройденное расстояние и время, проведенное в центре (40 x 40 см) и периферийных зонах.
Испытание в приподнятом крестообразном лабиринте.
Приподнятый крестообразный лабиринт, сделанный из белого акрилового пластика, состоял из четырех рукавов (каждое 28 × 5 см) и центральной области (5 × 5 см), возвышающейся на 50 см над полом.Два рукава были открыты, а два закрыты стенками высотой 15 см из того же материала. Мышей по отдельности помещали в центр лицом к открытой руке и позволяли исследовать в течение 5 мин. Оценивались следующие варианты поведения: время, проведенное в закрытых и открытых рукавах.
Тест в водном лабиринте Морриса.
Мышей помещали в круглый резервуар диаметром 120 см с водой комнатной температуры (22 ° C), который делали непрозрачным с помощью нетоксичной белой темперной краски. На стенах испытательной комнаты вокруг резервуара были размещены высококонтрастные черно-белые изображения, которые служили визуальными ориентирами.
На первом этапе теста мышей обучили находить видимую платформу диаметром 10 см, расположенную на высоте 1 см над водой. Если они не находили платформу в течение 60-секундного пробного лимита, их направляли к ней и позволяли оставаться в течение 20 секунд. После извлечения из бассейна мышей протирали полотенцами и помещали под нагревательную лампу для просушки. Было проведено пять испытаний, и местоположение платформы было изменено для каждого испытания.
На втором этапе мышей тестировали, чтобы найти скрытую платформу, которая была погружена на 1 см ниже поверхности в юго-восточном квадранте.Мышей тестировали один раз в день в течение четырех дней подряд во второй половине дня, пять испытаний на тренировке, по крайней мере, через 10 минут между испытаниями. Стартовые квадранты варьировались между испытаниями в одном и том же порядке для всех мышей. Задержка, необходимая для определения местоположения платформы, и длина их пути были записаны.
Испытание зонда проводили на 5 -й день , когда мыши плавали в течение 60 секунд без платформы в резервуаре. Регистрировалось время, проведенное в квадранте, где находилась затопленная платформа на предыдущих этапах.
Контекстно-зависимый тест на обусловленность страха.
Контекстуальное и управляемое условное обозначение страха проверяет способность мышей узнавать и запоминать связь между сигналами окружающей среды и отвращением. В этом тесте мышей помещали в камеру кондиционирования и давали порции условного стимула (CS) (слуховой сигнал) и отталкивающего безусловного стимула (US) (электрический разряд стопы). Камера кондиционирования состояла из непрозрачного акрилового пластика 30x30x21 см с прозрачной крышкой и противоударной сеткой на полу из металлических стержней диаметром 2 мм с промежутками 6 мм между проходами.Во время стадии кондиционирования в день 1 мышам позволяли свободно исследовать камеру в течение 120 секунд. После этого в течение 30 секунд подавали белый слуховой сигнал с шумом 55 дБ в виде CS, и в течение последних 2 секунд звука мышам давали удар ногой 0,8 мА в виде УЗИ. Презентация CS-US повторялась три раза за сеанс (120, 240 и 360 секунд после начала кондиционирования). После последнего удара ногой мышей оставляли в камерах в покое на 90 секунд.
После завершения сеанса кондиционирования мышей через 24 часа возвращали в ту же камеру кондиционирования и оценивали их поведение замирания для измерения контекстуально обусловленного страха (контекстный тест). Мышей помещали в камеру для кондиционирования, и им позволяли свободно исследовать камеру в течение 300 секунд без предъявления CS и US. Тест Cued был проведен в тот же день через два часа после контекстного теста. В этом испытании шокирующая сетка была удалена, а стены камеры были покрыты обоями с шахматным рисунком, что обеспечило новый контекст, не связанный с камерой кондиционирования.Мышей помещали в камеру для тестирования на 3 мин. В конце первых 3 минут слуховой сигнал CS, представленный во время кондиционирования, давался мышам на 3 минуты. Камеру кондиционирования страха протирали 70% спиртом после каждого теста. Память о страхе оценивалась на основе замораживания в условных сигналах или контекстах, которым мыши подвергались ранее. Переменными результата было время замораживания в контекстном тесте, а также в течение первых и последних 30 секунд теста с указанием.
Тест социального взаимодействия.
Два вида социального поведения (социальное взаимодействие и социальная память / распознавание новизны) были количественно оценены с помощью прямоугольного трехкамерного теста, который включал среднюю камеру размером 20x45x30 см, сделанную из акрилового пластика, с двумя отверстиями 10×10 см, ведущими к двум отдельным (левому и правому) камеры одинакового размера, каждая из которых содержит стальную клетку. Каждую мышь (экспериментальный субъект) помещали в среднюю камеру и позволяли исследовать и взаимодействовать в течение 10 минут, при этом правая камера была пустой, но с незнакомым сородичем (Stranger I) (контрольная мышь SPF того же пола, не являющаяся однопометником, содержащаяся в отдельный контейнер), удерживаемый в стальном кожухе в левой камере.Социальное взаимодействие определялось путем измерения времени, проведенного подопытным в камере с незнакомым сородичем, по сравнению с правой камерой, содержащей пустой корпус. Чтобы измерить социальную память (или узнавание новизны), новую мышь-стимул (Stranger II) поместили в ранее пустую правую камеру. Тестируемой мыши позволяли исследовать и взаимодействовать в течение 10 мин. Этапы теста следовали один за другим без промедления. Те же параметры, что и выше, были измерены для определения предпочтения испытуемого в отношении Незнакомца I или Незнакомца II.Социальную камеру протирали 70% спиртом после каждого теста.
Стереологическая оценка популяций нейронов и олигодендроцитов
После МРТ мозг извлекали из черепа, подвергали криозащите в 30% сахарозе и замораживали на сухом льду. Последовательные срезы толщиной 20 микрон разрезали на 400 микрон на криостате и помещали на предметные стекла, покрытые поли-L-лизином (Sigma-Aldrich, Сент-Луис, Миссури). Смежные серии использовали для окрашивания на маркеры нейронов и олигодендроцитов, а также на содержание миелина.
Срезы, используемые для иммуноокрашивания, блокировали 3% козьей сывороткой с последующей инкубацией с первичными антителами в течение ночи при 4 ° C. После этого следовала инкубация с биотинилированными вторичными антителами в течение 1 часа при 21 ° C и комплексом авидин-биотин (Vectastain Elite ABC Kit, Vector Laboratories, Burlingame, CA) в течение 1 часа. Цвет проявлялся с использованием 3,30-диаминобензидина (Sigma-Aldrich). В качестве первичных используемых антител были кроличьи анти-NeuN (ab177487, Abcam, 1: 400) и мышиные анти-Olig2 (MABN50, Millipore, 1: 400).Вторичными антителами были козьи анти-кроличьи ВА-1000 (1: 200) и козьи антимышиные ВА-9200 (1: 200), оба от Vector Laboratories. Меченые срезы визуализировали под микроскопом (Leica Microsystems, Wetzler), прикрепленным к моторизованному столику. Зонд оптического фракционатора в программном обеспечении StereoInvestigator (MBF Bioscience, Williston, VT) использовался для получения объективной оценки общего количества клеток или плотности клеток. Поскольку точные границы моторной коры трудно очертить точно, для оценки общего числа нейронов в этой области плотность нейронов в моторной коре оценивалась в 4 последовательных секциях, помеченных анти-NeuN из каждого образца мозга, начиная с 2 мм от передней части. головной мозг.Границы моторной коры были очерчены с помощью 5-кратного объектива, а подсчет клеток производился с помощью 40-кратного объектива. Размер рамки подсчета составлял 100 мкм х 100 мкм, а размер сетки составлял 600 мкм х 600 мкм. После завершения подсчета для каждого среза образца мозга рассчитывалась плотность клеток с использованием расчетной популяции и объема каждой подсчитанной площади, предоставленных программным обеспечением, и усреднялась для четырех срезов.
Чтобы оценить общее количество нейронов в гиппокампе, NeuN-положительный подсчет клеток выполнялся на всех серийных срезах, где присутствовал гиппокамп, обычно по 4–5 на мозг.Границы подразделения гиппокампа были тщательно определены с использованием атласа мозга мыши [39] и четкого морфологического признака заметных более мелких и более плотно организованных пирамидных нейронов CA1 по сравнению с относительно более крупными и менее упакованными нейронами, характеризующими CA2- Подразделения CA3 [40]. Границы коры областей CA1, CA2 / 3 и зубчатой извилины были очерчены с помощью 5-кратного объектива, а подсчет клеток производился с помощью 40-кратного объектива. Размер рамки счета составлял 50 мкм x 50 мкм, а размер сетки составлял 250 мкм x 250 мкм.Расчетное количество клеточной популяции рассчитывали по стереологической формуле.
Плотность олигодендроцитов оценивалась на одном срезе в StereoInvestigator, помещая счетную сетку с теми же параметрами, что и выше для NeuN, при 0 брегма для полосатого тела и областей коры, -0,94 брегмы для фимбрии и -1,28 брегмы для мозолистого тела и областей внутренней капсулы. . Плотность олигодендроцитов получали путем деления расчетной численности популяции на измеренный объем, предоставленный StereoInvestigator.
Статистический анализ
Данные представлены как среднее ± стандартная ошибка среднего. Сравнение групп лечения и пола для каждого возраста проводилось с помощью двухфакторного дисперсионного анализа. Сравнение групп после hoc и проводили методом Тьюки-Крамера. Скорректированное с поправкой на коэффициент ложного обнаружения (FDR) значение p (или значение q) также было рассчитано с использованием процедуры Бенджамини – Хохберга для корректировки множественных сравнений в объемных данных МРТ и данных MPF. Время задержки выхода на платформу в тесте водного лабиринта Морриса анализировали с помощью дисперсионного анализа с повторными измерениями (RM) с днем тренировки в качестве повторяющегося фактора и группой лечения в качестве фиксированного фактора.Для сравнения показателей результатов в поведенческих тестах использовалось заранее определенное ограниченное количество критериев исхода (<4), и никакой поправки на множественные сравнения не делалось [41]. Различия считались достоверными при α = 0,05.
Результаты
Магнитно-резонансная томография
Региональные изменения объема.
Региональные объемы головного мозга, полученные из in vivo, атласа, основанного на методологии парцелляции мозга, основанной на контракте ткани переноса намагниченности, суммированы в абсолютных единицах объема в таблице S1 и в процентах, нормализованных для мозга отдельной мыши, в таблице S2.Не было разницы в общем объеме мозга между мышами SPF и GF ни на четырех, ни на 12-недельном тестировании (рис. 2А). Относительный объем серого вещества был выше у мышей SPF в обонятельных луковицах, неокортексе и мозжечке (рис. 2B – 2D), но был ниже в таламусе (рис. 2E) и переднем мозге / перегородке (таблица S2) по сравнению с мышами GF через четыре недели. Размер обонятельных луковиц отличался в абсолютном объеме через 12 недель (таблица S1), и тест после hoc показал, что относительный размер обонятельных луковиц был значительно увеличен у самцов мышей GF (рис. 2В).В возрасте 12 недель самки мышей SPF имели значительно более высокий объем полосатого тела (F 1, 30 = 21,19, p <0,001), чем мыши GF при сравнении после hoc ( p = 0,0003, рис. 2F).
Рис. 2. Различия в региональном объеме мышей SPF и GF.
Региональные различия в объеме у мышей SPF и GF у четырех (n = 15 для самок и самцов мышей SPF; n = 6 для самок и n = 9 для самцов мышей GF) и 12 (n = 13 для самок и самцов SPF) и n = 7 для самок и самцов мышей GF) недельного возраста.Звездочки указывают на значимые различия, когда значение p составляет по крайней мере <0,05 между группами лечения или между отдельными группами в сравнениях после hoc , где фактор группы лечения двухфакторного дисперсионного анализа для каждого тестируемого возраста был значительным.
https://doi.org/10.1371/journal.pone.0201829.g002
Локальные изменения объема головного мозга по данным тензорной морфометрии.
Тензорная морфометрия (TBM) — это метод статистического картирования, основанный на значении детерминанта Якоби с цветовой кодировкой, разработанный для количественной оценки региональных структурных различий относительно соответствующего анатомического шаблона [42, 43].Не было обнаружено значительных различий между группами через четыре недели. Значительное региональное расширение мозга у мышей GF было обнаружено в обонятельных луковицах и префронтальной коре через 12 недель (рис. 3).
Рис. 3. Сравнение тензорной морфометрии мышей SPF и GF.
Количество животных: мыши SPF (n = 5 как для самок, так и для самцов) и GF (n = 5 для самок и самцов) в возрасте 12 недель. Не было обнаружено значительных различий в возрасте четырех недель с одинаковым количеством животных в каждой группе.Псевдоцветная статистическая параметрическая карта накладывается на шаблон шкалы серого. Статистически значимое расширение регионального объема в обонятельных луковицах и префронтальной коре в группе GF показано псевдоцветными вокселями в красно-желтой шкале. Цветная полоса показывает скорректированный диапазон значений p , а значение p <0,05 считалось статистически значимым.
https://doi.org/10.1371/journal.pone.0201829.g003
Организация белого вещества и миелинизация, на которую указывает фракционная анизотропия.Мыши
SPF имели значительно увеличенную фракционную анизотропию в фимбриях, передней спайке, мозолистом теле, зрительном тракте, внутренней капсуле и перивентрикулярном белом веществе через четыре недели тестирования (рис. 4). Не было обнаружено значительных различий между мышами SPF и GF через 12 недель.
Рис. 4. Статистические параметрические карты сравнения мышей SPF и GF с использованием показателей FA и радиальной диффузии.
Количество животных: мыши SPF (n = 5 как для самок, так и для самцов) и GF (n = 5 для самок и самцов) через четыре недели.Не было обнаружено значительных различий в возрасте 12 недель с одинаковым количеством животных в каждой группе. Зеленые линии скелета были наложены на шкалу серого FA, показанную на сагиттальном, корональном и горизонтальном сечениях. Статистически значимые воксели, у которых параметры были меньше в группе GF, изображены в красно-желтой шкале для FA. Цветная полоса указывает диапазон скорректированных значений p , а значение p <0,05 считалось статистически значимым. Сокращения анатомических структур: ac-передняя комиссура, CC-мозолистое тело, ic-внутренняя капсула, фимбрия, отозрительный тракт, pwm-перивентрикулярное белое вещество.
https://doi.org/10.1371/journal.pone.0201829.g004
Визуализация фракции макромолекулярных протонов (биомаркер миелинизации).
Значения групп MPF приведены в таблице S3. У самцов мышей SPF через четыре недели наблюдалось повышение MPF в общем мозге (F 1,38 = 6,51, p = 0,015), в белом веществе, включая мозолистое тело, переднюю комиссуру и внутреннюю капсулу (рис. 5A – 5D), так как хорошо в структурах серого вещества, включая неокортекс (рис. 5E), гиппокамп (рис. 5F), гипоталамус и ствол мозга / средний мозг (таблица S3).Гендерный фактор и факторы взаимодействия «пол x группа» не были значимыми через четыре недели. Через 12 недель тестирования MPF у самцов SPF был выше во внутренней капсуле (рис. 5D), а гендерный фактор был значимым для всех разделенных областей (таблица S3).
Рис. 5. Доля протонов макромолекул (в процентах) у мышей SPF и GF.
Количество животных — четыре (n = 15 для самок и самцов мышей SPF; n = 6 для самок и n = 9 для самцов мышей GF) и 12 (n = 13 для самок и самцов SPF и n = 7 для обоих самок и самцов мышей GF) недельного возраста.Звездочки указывают на значимые различия, когда значение p составляет по меньшей мере <0,05 между группами в апостериорных сравнениях , где фактор группы лечения двухфакторного дисперсионного анализа для каждого тестируемого возраста был значительным.
https://doi.org/10.1371/journal.pone.0201829.g005
Содержание миелина при окрашивании Luxol Fast blue
Интенсивность окрашивания LFB была значительно выше у мышей SPF по сравнению с мышами GF (оба пола вместе) в передней спайке, мозолистом теле и внутренней капсуле в возрасте четырех недель (рис. 6A) и во внутренней капсуле в возрасте 12 недель ( Рис 6B).
Рис. 6. Определение содержания миелина с помощью окрашивания Luxol Fast blue.
Количество животных в возрасте четырех (n = 6 для мышей SPF и GF) (A) и 12 (n = 6 для мышей SPF и GF) возраста (B). Звездочки указывают на значительные различия, когда значение p составляет по крайней мере <0,05 между группами SPF и GF.
https://doi.org/10.1371/journal.pone.0201829.g006
Поведенческие тесты
Испытания в открытом поле и в приподнятом крестообразном лабиринте.
Влияние микробиоты на общую двигательную активность нейронов и тревожное поведение оценивали с использованием теста открытого поля (OFT) и приподнятого крестообразного лабиринта (EPM).Ключевыми макроструктурами мозга, которые участвуют в нервных цепях тревоги, являются миндалевидное тело, ядро ложа терминальной полоски, вентральный гиппокамп и префронтальная кора, но не ограничиваются ими. Мыши SPF были значительно более мобильными (рис. 7A, F 1,39 = 16,85, p <0,001, двусторонний дисперсионный анализ) и имели более высокую среднюю скорость (рис. 7B, F 1,39 = 14,95, p <0,001, двухфакторный дисперсионный анализ), чем у мышей GF через четыре недели в OFT. Не было обнаружено различий в гендерном факторе и взаимодействии микробиоты и пола через четыре недели.При сравнении после hoc, четырехнедельные самки и самцы SPF имели значительно более высокое мобильное время ( p = 0,002 и p = 0,043, соответственно) и среднюю скорость ( p = 0,0049 и p = 0,016 соответственно), чем самок и самцов GF. В результате мыши SPF прошли значительно большее расстояние (48,40 ± 2,49 м), чем мыши GF (30,72 ± 2,06 м) за четыре недели (F 1, 39 = 15,37, p <0,001, двусторонний дисперсионный анализ ANOVA). Не было обнаружено различий в гендерном факторе и взаимодействии микробиоты и пола в расстоянии путешествия в возрасте четырех недель.При сравнении после hoc самки и самцы SPF в возрасте четырех недель значительно отличались от самок и самцов GF ( p = 0,0048 и p = 0,018, соответственно). Кроме того, четырехнедельные мыши SPF проводили значительно больше времени в центральной области (рис. 7C, F 1, 39 = 4,26, p = 0,045, двусторонний дисперсионный анализ ANOVA). Никакой разницы во времени передвижения, средней скорости, пройденном расстоянии или времени, проведенном в центральной зоне, не было обнаружено при 12-недельном тестировании. В тесте EPM не было обнаружено различий во времени, проведенном в закрытых руках, в зависимости от пола, микробиоты или микробиоты x гендерного взаимодействия в возрасте четырех или 12 недель (рис. 7D).
Рис. 7. Поведенческие тесты в открытом поле и приподнятом крестообразном лабиринте.
Мыши SPFбыли более мобильными (A) и передвигались с большей скоростью (B) через четыре недели (n = 15 для самок и самцов мышей SPF; n = 6 для самок и n = 9 для самцов мышей GF), но не через 12 недель (n = 13 как для самок, так и для самцов SPF и n = 7 для самок и самцов мышей GF). Мыши SPF проводили больше времени в центральном квадранте в тесте «открытое поле» через четыре недели, но не через 12 недель (C). D. GF мыши не отличались от мышей SPF через четыре недели и через 12 недель по времени, проведенному в закрытой руке в приподнятом крестообразном лабиринте.Звездочки указывают на значительные различия, когда значение p составляет по крайней мере <0,05.
https://doi.org/10.1371/journal.pone.0201829.g007
Водный лабиринт Морриса.
Пространственное обучение и память, которые тесно связаны со схемой гиппокампа, а также префронтальной корой, поясной извилиной корой и неостриатумом [44], оценивались с использованием водного лабиринта Морриса у мышей SPF и GF. При тестировании задержки для поиска платформы во время тренировочных дней с помощью двухфакторного дисперсионного анализа с одним повторным измерением (этапом) фактор тренировочного дня был значимым на обеих четырех неделях (F 3, 102 = 11.72, p <0,001) и 12 недель (F 2, 102 = 4,8, p <0,001), демонстрируя, что как в группах SPF, так и в группах GF время задержки для поиска скрытой платформы уменьшилось во время тренировки и в обеих группах. обучались (рис. 8A и 8B). Фактор микробиоты существенно не отличался через четыре недели, но был значимым через 12 недель (F 1, 36 = 11,72, p = 0,035) во время обучения между мышами SPF и GF. Анализ после hoc показал, что у самок SPF была более короткая латентность, чтобы найти платформу в конце тренировки (день 4) (F 1, 16 = 12.09, p = 0,003).
Рис. 8. Поведенческие тесты памяти и обучения с использованием водного лабиринта Морриса и кондиционирования страха.
Не было обнаружено значительной разницы во время обучения между мышами SPF (n = 15 для самок и самцов) и GF (n = 6 для самок и n = 9 для самцов) через 4 недели (A), но через 12 недель (B). ) Мыши SPF (n = 13 как для самок, так и для самцов) имели значительно более высокий наклон кривой обучения, чем мыши GF (n = 7 как для самок, так и для самцов), по данным дисперсионного анализа RM ANOVA. (С). Мыши SPF проводили значительно больше времени в квадранте платформы во время зондирования теста водного лабиринта Морриса через 12 недель, но не отличались от мышей SPF через 4 недели.(D). Скорость плавания самцов SPF была значительно выше, чем у самцов GF через 12 недель. (E). Время замирания было больше в тесте условного рефлекса страха у самцов мышей SPF через 12 недель. (F). Время замораживания было короче для мышей SPF во второй части теста условного рефлекса страха (представлены разные камеры и звуковые сигналы) на 12 неделе. Звездочки указывают на значительные различия, когда значение p составляет по крайней мере <0,05.
https://doi.org/10.1371/journal.pone.0201829.g008
Испытание зонда, в котором платформа была удалена после этапов обучения, не выявило значительной разницы между мышами SPF и GF во времени, проведенном в квадранте на четырехнедельная временная точка, ни для мужчин, ни для женщин (рис. 8C).Через 12 недель тестирования мыши SPF проводили значительно больше времени в квадранте, чем мыши GF (Рис. 8C, F 1, 36 = 11,76, p = 0,0015, двусторонний дисперсионный анализ ANOVA) и у обоих полов по post- hoc сравнения.
Никакой разницы в средней скорости не было обнаружено за четыре недели тестирования (рис. 8D). В возрасте 12 недель мыши SPF имели значительно более высокую среднюю скорость плавания, чем у мышей GF (F 1, 36 = 11,72, p = 0,0016, двусторонний дисперсионный анализ ANOVA). При сравнении после и у 12-недельных самцов SPF была более высокая средняя скорость, чем у самцов GF (рис. 8D, p = 0.002).
Испытание на формирование страха.
Для характеристики зависимой от гиппокампа и / или миндалины долговременной памяти у мышей SPF и GF мышей подвергали контекстуальной / управляемой условной рефлексией страха. Не было обнаружено разницы во времени замораживания между мышами SPF и GF любого пола до кондиционирования. Не было разницы во времени замораживания между мышами SPF и GF при условном условном рефлексе страха (измененная среда и отсутствие акустического представления CS) при четырехнедельном тестировании (рис. 8E).Однако у SPF было значительно более длительное время замораживания, чем у мышей GF через 12 недель (F 1, 39 = 11,44, p = 0,022), причем у самцов мышей SPF время замораживания было значительно больше (после теста hoc , p = 0,032). Гендерный фактор был значимым (F 1, 39 = 11,44, p = 0,002) из-за значительно более короткого времени замораживания у самцов SPF ( p = 0,047) в возрасте четырех недель. В возрасте 12 недель время замораживания мышей SPF на этой стадии было значительно дольше, чем у мышей GF (F 1, 39 = 11.44, p = 0,002). Тест после hoc показал, что время замораживания у самцов SPF было значительно дольше, чем у самцов GF ( p = 0,047). Никакой разницы во времени замораживания во второй части теста условного рефлекса страха (измененная среда и акустическая презентация CS) не было обнаружено в возрасте 4 недель, но в возрасте 12 недель время замораживания мышей SPF было значительно короче, чем у мышей GF. (F 1,39 = 9,28, p = 0,0043, рис. 8F). Тест после hoc показал, что время замораживания SPF было значительно короче, чем у мышей GF ( p = 0.047) у обоих полов.
Тесты социального взаимодействия: коммуникабельность и новизна.
Мы дополнительно исследовали, существует ли разница между мышами SPF и GF в социальных взаимодействиях в трехкамерном социальном тесте, отражающем в основном префронтальную кору и пластичность миелинизации. В тесте на общительность для предпочтения социальной и пустой клетки не было обнаружено значительных различий между мышами SPF и GF по времени, проведенному в камере с мышами (Stranger 1, рис. 9A) или с пустой клеткой, ни в возрасте четырех, ни в 12 недель тестирования ( Фиг.9B, с индексом дискриминации, показанным на рисунке A в S1 Fig).В тесте на социальную новизну не было обнаружено различий между группами SPF и GF во времени, проведенном со знакомой мышью (рис. 9C), но мыши SPF проводили значительно больше времени с новой мышью (Stranger 2), чем мыши GF у обоих полов. 12 недель (F 1,32 = 5,10, p = 0,030) (рис. 9D, с индексом дискриминации, показанным на рис. B на рис. S1).
Рис. 9. Тесты социального взаимодействия с использованием трехкомнатной камеры.
В тесте на общительность мыши SPF и GF не различались по времени, проведенному в камере со странной мышью (A) или с пустой клеткой (B) у четырех (n = 15 как для самок, так и для самцов мышей SPF; n = 6 для самок и n = 9 для самцов мышей GF) или 12 недель (n = 13 для самок и самцов SPF и n = 7 для самок и самцов мышей GF).В тесте на социальную новизну не было обнаружено различий между группами SPF и GF по времени, проведенному со знакомой мышью (C), но наблюдалось значительное уменьшение времени, проведенного с новой мышью у мышей GF через 12 недель (D). Звездочки указывают на значительные различия, когда значение p составляет по крайней мере <0,05.
https://doi.org/10.1371/journal.pone.0201829.g009
Стереологическая оценка числа нейронов и олигодендроцитов
Пытаясь объяснить разницу в локомоции через четыре недели и в пространственной памяти через 12 недель, мы оценили количество нейронов в моторной коре и гиппокампе, а также количество олигодендроцитов в коре и нескольких областях белого вещества.Количество нейронов, идентифицированных ядерным антигеном нейронов (NeuN), оценивали для всего гиппокампа. Не было обнаружено различий между мышами SPF и GF ни в CA1, CA2 / 3, ни в зубчатой извилине гиппокампа через 12 недель (фиг. 10A). Поскольку точные границы моторной коры определить неоднозначно, плотность нейронов оценивалась в центральной части моторной коры. Не было обнаружено значительных различий в плотности нейронов между мышами SPF и GF в возрасте четырех недель (фиг. 10B). Плотность олигодендроцитов, определяемая маркером Olig2 линии олигодендроцитов, была значительно выше в мозолистом теле мышей SPF через 12 недель ( p = 0.018) (рис. 10C), с тенденцией к увеличению внутренней капсулы, фимбрии, коры и полосатого тела, которые не достигли значительной разницы.
Рис. 10. Стереологическая оценка количества клеток между мышами SPF и GF.
(А). Оценка количества нейронов в областях гиппокампа в возрасте 12 недель (n = 6 для мышей SPF и GF). (В). Плотность нейронов в моторной коре через четыре недели. (С). Плотность олигодендроцитов в областях белого и серого вещества в возрасте 12 недель. Звездочки указывают на значительные различия, когда значение p по крайней мере <0.05.
https://doi.org/10.1371/journal.pone.0201829.g010
Обсуждение
Настоящее исследование вносит вклад в быстро растущий объем литературы, посвященной роли кишечной микробиоты и взаимодействий между кишечником и мозгом в формировании развития и поведения мозга. Новые аспекты исследования заключались в применении комбинации методов фокусировки функций (расширенная МРТ нейровизуализация и поведенческие тесты) с дополнительными иммуногистологическими исследованиями для изучения влияния комменсальных бактерий на структуру мозга и поведение мышей C57BL / 6J.Кроме того, новое количественное МРТ-картирование фракции макромолекул было использовано для оценки влияния комменсальных бактерий на миелинизацию во многих структурах мозга. МРТ in vivo, и ex vivo, показала, что мыши SPF с комменсальными бактериями демонстрировали несколько более крупных областей в сером веществе и других областях мозга, чем мыши GF без комменсальных бактерий. Мы поспособствовали тому, что наблюдаемые мыши SPF были более миелинизированы в структурах серого вещества, включая неокортекс, гиппокамп и несколько основных трактов белого вещества у молодых самцов мышей в возрасте четырех недель и во внутренней капсуле у самцов в возрасте 12 недель.Эти различия в миелинизации были подтверждены с помощью быстрого окрашивания Luxol blue на миелин. Что еще более важно, наблюдаемые различия в структуре мозга коррелировали с различиями в поведении мышей SPF и GF (рис. 11). Мы продемонстрировали, что мыши SPF и GF демонстрируют временные различия в поведении, связанном с тревогой, и долгосрочные различия в пространственной памяти, контекстной и управляемой памяти, а также в социальной новизне. Объемные и организационные различия между мышами SPF и GF, наблюдаемые в развитии фимбрии, мозолистого тела, внутренней капсулы и дорсального полосатого тела, могут объяснить разницу в двигательных и когнитивных результатах в открытом поле, водном лабиринте Морриса и тестах на кондиционирование страхом; тогда как различия в объеме и миелинизации в неокортексе и префронтальной коре могут способствовать различию в социальном поведении.Таким образом, связывая пораженные определенные области мозга с соответствующими функциями, наше исследование предоставляет доказательства того, что комменсальные бактерии могут влиять на развитие и поведение мозга.
Во-первых, наше исследование показало, что микробиота влияет на развитие объемов различных структур мозга. МРТ широко используется в исследованиях неврологического развития, поскольку с ее помощью можно неинвазивно количественно оценить структурные изменения мозга и коррелировать с моторным и когнитивным развитием в клинических испытаниях [11] и исследованиях на животных [13, 45].Исследования нормального созревания мозга у людей сообщают об объемных изменениях серого и белого вещества в период между детством и подростковым возрастом, но относительно стабильным общим объемом мозга в этот период [14, 46]. Точно так же у мышей общий объем мозга при МРТ достигает стабильности через три недели, но созревание серого и белого вещества, включая миелинизацию, непрерывно прогрессирует в течение первых трех месяцев [47]. В нашем исследовании, насколько нам известно, впервые систематически изучалась роль микробиоты в объемных различиях.Использовались как автоматическая парцелляция мозга in vivo , так и морфометрия на основе тензора ex vivo MRI. Различия наблюдались в основном в ранний период после отъема. У мышей SPF временно увеличился объем коры, мозжечка, обонятельных луковиц и уменьшился объем таламуса и объема мозга / перегородки по сравнению с мышами GF в возрасте четырех недель. Эти наблюдения согласуются с мнением [48], что ранняя микробиота кишечника важна для раннего развития мозга.
В нашем следующем исследовании изучалось влияние микробиоты на развитие белого вещества.DTI — это часто используемый метод нейровизуализации для оценки целостности белого вещества и использует производный индекс фракционной анизотропии (FA) в качестве суррогатной меры для организации белого вещества и миелинизации. Изменения значений FA могут оценивать созревание белого вещества, отражая изменения числа аксонов, целостности структуры миелина и целостности аксонального цитоскелета [49]. В нашем исследовании мы обнаружили значительно более высокие значения FA в фимбриях, мозолистом теле, внутренней капсуле, перивентрикулярном белом веществе и зрительных трактах как у самцов, так и у самок мышей SPF через четыре недели по сравнению с мышами GF.Большое количество литературы связывает различия в FA с поведенческими исходами. В исследованиях на животных, поражения фимбрии-свода (края гиппокампа) изменяли пространственное обучение / память и локомоцию [50, 51]. В исследованиях на людях было показано, что значения FA нескольких участков белого вещества, включая фимбрию и мозолистое тело, положительно коррелируют с мелкомоторными и когнитивными баллами по шкале Бейли развития младенцев и малышей у детей, рожденных недоношенными и изученных в двухлетнем исправленном возрасте [52 ].Мозолистое тело координирует сенсорные, моторные, когнитивные и эмоциональные функции обоих полушарий у младенцев [53]. Показано, что значения FA мозолистого тела положительно связаны с когнитивными результатами [54–56]. Значения FA внутренней капсулы [57] связаны с лучшими связями и моторикой у подростков, рожденных недоношенными [58], и когнитивными уровнями у пациентов с перивентрикулярной лейкомаляцией [54] [59, 60]. Значения FA перивентрикулярного белого вещества положительно коррелируют с развитием белого вещества и когнитивными функциями в нормальной педиатрической популяции [61] и могут прогнозировать повреждение белого вещества при некоторых состояниях, связанных с недоношенностью [58].В соответствии с этой литературой, зависимые от микробиоты различия, которые мы обнаруживаем в значениях FA фимбрии, мозолистого тела, внутренней капсулы и перивентрикулярного белого вещества, могут объяснить различия в двигательной активности в тесте открытого поля, пространственной памяти в водном лабиринте Морриса и контекстной памяти в в нашем исследовании был проведен тест на обусловленность страха.
Одним из основных результатов этого исследования является ранее не описанная микробиотозависимая гипомиелинизация нескольких структур серого вещества, включая неокортекс, гиппокамп, ствол мозга и основные тракты белого вещества, включая мозолистое тело, переднюю спайку и внутреннюю капсулу, особенно у мышей GF с использованием MPF. изображения.Миелинизация головного мозга и белого вещества обеспечивает оценку созревания мозга [18, 62, 63], а состояние миелинизации мозга является сильным показателем постнатальной неврологической функциональной зрелости [64]. В нашем исследовании разница была наиболее значительной через четыре недели, но не через 12 недель, за исключением внутренней капсулы. Окрашивание LFB соответствовало данным MPF в большинстве областей белого вещества, в частности, подтверждая более высокую степень миелинизации у мышей SPF по сравнению с мышами GF во внутренней капсуле через 12 недель.Обнаружение кортикальной гипомиелинизации через четыре недели в нашем исследовании у мышей GF контрастирует с ранее описанной гипермиелинизацией префронтальной коры, обнаруженной через 10 недель у мышей Swiss Webster [21]. Префронтальная кора объединяет внешние стимулы и контролирует несколько областей сложного поведения [65]. Мы не обнаружили различий между мышами SPF и GF в миелинизации, особенно в префронтальной коре, с использованием МРТ и окрашивания LFB через 12 недель. Расхождения между нашими и другими выводами можно объяснить различными линиями мышей и методами, используемыми для оценки миелинизации.Использование MPF при окрашивании МРТ и LFB, оба из которых являются методами визуализации, в нашем исследовании обеспечивает пространственную специфичность отбора проб серого и белого вещества. Нарушения миелинизации у молодых животных GF предполагают важность комменсальной микробиоты для миелинизации мозга и расширяют влияние ранней микробной колонизации на созревание мозга.
Было проведено несколько исследований, в которых выявлены различия в поведении мышей SPF и GF, особенно в отношении избегания риска и исследовательских задач, обучения и памяти, а также социального поведения [4].Однако текущие литературные данные не согласуются с поведением у разных линий мышей. На сегодняшний день в линиях мышей BALB / c, NMRI и Swiss Webster сообщалось о снижении исследовательского поведения у мышей SPF с использованием тестов открытого поля и приподнятого лабиринта [6, 8, 20, 66], в то время как мыши C57BL / 6N SPF проявляли повышенную тревогу в Тест предпочтения свет / темнота, но снижение тревожности в тесте с понижением по сравнению с мышами GF [67]. Даже в трех подгруппах мышей C57BL / 6 мыши C57BL / 6J, C57 / BL6N и C57BL / 6C, C57BL / 6J показали наиболее исследовательское поведение в открытом поле и тест «приподнятый крестообразный лабиринт» среди трех подгрупп [68] .В наших исследованиях мы сообщили в тесте «открытое поле», что мыши SPF линии C57BL / 6J проявляли более высокую исследовательскую активность и, таким образом, более выраженное анксиолитическое поведение и двигательную активность в возрасте четырех недель по сравнению с мышами GF. Мы предполагаем, что различия в этих исследованиях, включая наше, можно объяснить различиями в генетическом фоне и профилях микробиома у линий мышей. Эти результаты отражают сложность двунаправленных взаимодействий микробиоты и хозяина, одновременно указывая на то, что микробиота действительно играет роль в формировании исследовательского / тревожного поведения хозяина.
Ранее снижение рабочей памяти, тестируемое с помощью распознавания новых объектов, было обнаружено у самок мышей GF Swiss Webster [69] и мышей SPF C57BL / 6, получавших лечение антибиотиками [70]. Наши данные также показывают, что мыши SPF имеют лучшую пространственную память в возрасте 12 недель в тесте в водном лабиринте Морриса по сравнению с мышами GF. Ранние исследования показали, что поражения полосатого тела ухудшают способность животных выполнять задачи пространственного обучения [71–73]. Принимая во внимание роль дорсомедиального полосатого тела в пространственно-когнитивной функции, уменьшение объема полосатого тела, описанное в этом исследовании, может объяснить дефицит пространственной памяти у мышей GF в водном лабиринте Морриса в возрасте 12 недель.Мы также демонстрируем, что самцы мышей SPF обладают лучшей контекстной памятью в тесте условного рефлекса страха в возрасте 12 недель, который зависит от гиппокампа и лобной / вентромедиальной / поясной коры головного мозга [74]. Однако у них снижена задержка запоминания по сравнению с мышами GF, которая считается зависимой от миндалины [75]. Недавнее исследование Hoban et . и . Использование C57BL / 6N продемонстрировало, что мыши SPF лучше удерживали память, но не отличались от мышей GF по контекстному воспроизведению [76].Расхождение между этими данными можно объяснить разными линиями мышей. Исследования показали, что у мышей C57BL / 6N контекстная память лучше, чем у мышей C57BL / 6J, но не было обнаружено различий в памяти с указанием между двумя подтипами [77]. Различия, наблюдаемые в исследованиях, также могут быть связаны с применяемыми протоколами экспериментов. Мы использовали традиционный протокол 24-часового удержания, в то время как Hoban и . и . использовали модифицированный протокол шестичасового удерживания.
Социальные нарушения у лиц с несколькими неврологическими расстройствами, такими как гиперактивное расстройство с дефицитом внимания [78], аутизм [79], расстройство поведения [80] и компульсивное обсессивное расстройство [81], были зарегистрированы у детей и во взрослом возрасте.Интересно, что эти расстройства обычно связаны с проблемами желудочно-кишечного тракта и дисбактериозом кишечной микробиоты [82, 83]. Мы не обнаружили разницы в общительности между мышами C57BL / 6J SPF и GF. Однако мыши SPF лучше отдавали предпочтение социальной новизне, чем мыши GF, с более выраженными эффектами, обнаруженными у самцов. Предыдущие исследования с использованием мышей Swiss Webster дали противоречивые результаты в отношении общительности и социальной новизны у мышей SPF по сравнению с мышами GF [10] [84]. Считалось, что расхождения между предыдущими исследованиями отчасти связаны с различиями в протоколах, в частности с использованием разных линий социальных партнеров и возрастом животных при тестировании.
Взятые вместе, различные проявления поведения в этих исследованиях, включая наше, можно объяснить различиями в генетическом фоне, а также различиями в микробиоте разных линий мышей [67, 85–87]. Другими потенциальными факторами, влияющими на результаты, являются возраст животных на момент проведения тестов, а также выбранные тесты и протоколы [4]. Поэтому мы утверждаем, что интерпретация этих исследований должна уделять особое внимание этим факторам. Срочно необходимы дальнейшие исследования механизмов, с помощью которых микробиота кишечника взаимодействует с мозгом.
Имеется хорошо документированный половой диморфизм поведения в клинических исследованиях и исследованиях на животных [88–91]. У людей распространенность депрессии или тревоги в два-четыре раза выше у женщин, тогда как аутизм и СДВГ чаще встречаются у мужчин [88, 92]. Что касается когнитивных функций, женщины, кажется, лучше справляются с кратковременной памятью, чем мужчины, тогда как мужчины лучше справляются с пространственными задачами [93–95]. Ранее сообщалось, что у животных самки мышей C57BL / 6J более тревожны в тесте EPM, чем самцы, но не было обнаружено различий между полами в исследовательской деятельности в тесте «открытое поле» [90].Также не было обнаружено различий в общительности и новизне между самками и самцами мышей C57BL / 6J в трехкамерном тесте [91]. В тесте Морриса в водном лабиринте самцы крыс демонстрируют преимущества в пространственном обучении, зависящем от гиппокампа, и эталонной памяти для крыс независимо от линий, протоколов, возраста и условий выращивания, но мыши не показывают половых различий [96, 97]. Наши данные в значительной степени согласуются с большинством опубликованных исследований на мышах C57BL / 6J, в которых мы не обнаружили значительных различий между полами (рис. 7–9).Это требует осторожности при переводе данных моделей животных в поведение человека при использовании определенных штаммов грызунов. Для изучения полового диморфизма требуются более тщательно спланированные и контролируемые эксперименты на животных, поскольку многие факторы, такие как генетическая связь, гормональная стадия, циркадные ритмы и различные реакции самок и самцов животных на режим тренировки, могут влиять на поведение грызунов. Наши результаты, однако, укрепляют представление о том, что различия в поведении, которые мы наблюдали в этом исследовании, были вызваны присутствием или отсутствием микробиоты.Необходимо продолжить изучение того, как присутствие различных микробных сообществ или отсутствие микробиоты по-разному влияет на пол. Поскольку большинство наблюдаемых нами поведений не демонстрирует половой диморфизм, мы специально обнаружили, что самки мышей GF имели дефицит пространственного обучения, а самцы мышей GF имели дефицит контекстной памяти. В одном из недавних ограниченных исследований [98] колонизация мышей GF в раннем постнатальном периоде с помощью человеческих Bifidobacterium spp. улучшила распознавающую память как у самок, так и у самцов мышей, но восстановила тревожное поведение только у самок мышей GF.Механизмы этого специфичного для пола поведения хозяина и взаимодействия микробиоты необходимы для перевода текущих результатов в клиническое применение.
В совокупности новые доклинические исследования сделали микробиоту потенциальной мишенью для улучшения пластичности и функций мозга. Текущие экспериментальные подходы к исследованию влияния микробиоты на функции мозга включают использование пребиотиков [99–101], пробиотиков [98, 102, 103] или антибиотиков [23, 70, 104, 105] для управления микробиотой кишечника, а также « гуманизированные »модели свободных от микробов мышей [9, 98, 106].Например, олигосахариды грудного молока 3′-сиалиллактоза или 6′-сиалиллактоза в качестве пребиотика нормализовали вызванное стрессом тревожное поведение [101]. Кормление Bifidobacteria breve 1205 врожденно тревожным мышам BALB / c уменьшало общее тревожное поведение в тесте EPM [102]. Нарушение микробиоты кишечника пероральным приемом антибиотиков вызывало депрессивное поведение в тесте подвешивания за хвост и снижало социальную новизну в трехкамерном тесте социального взаимодействия у мышей C57BL / 6J [104].Хотя в этих исследованиях изучались изменения поведения и молекулярные изменения в мозге из-за манипуляций с микробиотой, знания о затронутой структуре и организации мозга отсутствовали. Наши результаты описывают фундаментальные различия в структурах мозга и поведении мышей SPF и GF, чтобы заложить основу для будущих исследований по выявлению и потенциальному нацеливанию структур и / или функций мозга, представляющих интерес для терапевтического развития.
Ограничением исследования было то, что, хотя наши животные были размещены в изокажах с положительным давлением, которые, как было показано в нескольких исследованиях, обладают способностью сохранять идентичность микробных сообществ у мышей GF в течение шести месяцев [107, 108], поэтому исключая возможное заражение во время транспортировки и содержания, мы не поддерживали статус свободных от микробиоты мышей GF в течение девятидневного поведенческого тестирования.Таким образом, мы называем этих мышей «ex-GF». Маловероятно, что морфологические изменения произойдут в течение короткого периода тестирования в девять дней после того, как мыши GF впервые столкнулись с окружающей средой в нашем исследовании. Другие исследования показали, что уровни тревожности [67], нарушение социального познания [10] и стрессовая реакция HPA [9] не были затронуты во время стандартизации взрослых мышей GF и у колонизированных мышей GF после отъема, временных точек, используемых в этом исследовании. Кроме того, мышей SPF в нашем исследовании подвергали той же «новой» среде, что и мышей GF.Любые эффекты из-за этого короткого знакомства с новым микробным сообществом должны применяться как к мышам SPF, так и к GF, а не только к GF. Таким образом, наши результаты в развитии мозга и различиях в поведении мышей SPF и ex-GF все еще можно интерпретировать как эффект комменсальной микробиоты.
В совокупности настоящее исследование предоставляет убедительные доказательства в поддержку идеи о том, что микробиота влияет на объем мозга, развитие белого вещества и миелинизацию, а также на тревожность, когнитивные способности, память и социальные функции.Различия, наблюдаемые через четыре недели, предполагают влияние микробиоты именно на раннее развитие мозга. Может существовать период развития, когда влияние кишечной микробиоты на развитие мозга является наибольшим, а в более позднем возрасте структурно не обнаруживается. Это подчеркивает важность дальнейших визуализационных и поведенческих исследований влияния микробиоты кишечника на раннее развитие мозга.
Система Каратэ Дробышевского: БАССАЙ ДАИ Часть 1 Куро Оби Бой 1 Сантори Бункай Кумитэ :: Медиа :: Видео о боевых искусствах FightCon
Бункай Бассай Дай в действии! Бассай Камаэ для Hand Grab Defense и Qinna Armlock.Моротэ Ути Уке для защиты ножом или бутылкой, разоружения и уничтожения оружия. Ути Уке и Гьяку Ути Уке Комбинация для защиты ножом или бутылкой, блокировки руки, обезоруживания оружия и Атеми.
Есть много вариантов бункай Бассай Дай ката. Мы показываем вам одну из последних разработок. Это настоящий Дан-тест, который этим летом провела Алина Чепулионис. Экзамен длился два часа, и она смогла продемонстрировать полный бункай Бассай Дай в форме боя черного пояса против нескольких вооруженных противников.
Революция нашего бункая состоит в том, что мы разделили Бассаи Дай на несколько отдельных сражений против нескольких вооруженных противников. Мы называем их «Бой Куро Оби» или «Бой за черный пояс». В каждом из этих боев нападающие идут по разным путям атаки: только спереди, спереди и сзади, сбоку, спереди и сзади и т. Д. Например, в первом бою все три противника на одной стороне и одновременно во втором бою. они фасадные и боковые фасады и т. д.
В ходе демонстрации ката варианты штурмовых бункаев и оружия постоянно меняются.Мы работаем над повышением боевого потенциала каждой новой версии бункая. Другими словами, все, даже самые примитивные движения ката, мы используем для победы над противником. Бункай ката должно соответствовать девизу старейшего каратэ «Иккен Хиссацу» (一拳 必殺) или «Уничтожить одним ударом». Идеальный бункай: один ход — один победить врага! Мы смогли применить первые движения ката Бассаи Дай в битве против трех противников, действуя в соответствии с древним девизом «Иккен Хиссацу». Этот бункай мы называем Сунтори Бункай Кумитэ.Для борьбы здесь применяют все движения, даже первое камаэ ката.
Выполняя на практике эти комбинации, вы должны помнить все советы сенсеев из Японии. На всех семинарах каждый раз акцентируют внимание на важности работы тазобедренного сустава в карате: акцент на фундаментальном коси-но кайтен (вращение бедра). Без этой работы успешная реализация данной комбинации невозможна!
ВНИМАНИЕ: Практикуйтесь на свой страх и риск! Ничто из этого не является юридической консультацией, вы и только вы несете ответственность за свои действия и выбор, который вы делаете!
В этом видео мы представляем бункай следующих элементов ката Бассай Дай:
1.Басай-камаэ. Хейсоку-дачи.
2. Migi Te Morote Uke. Коса-дачи.
3. Маватте, Хидари Учи-укэ. Хидари-Дзенкуцу-дачи.
4. Гьяку-ханми, Миги Гьяку Учи -укэ. Хидари-Дзенкуцу-дачи.
Бункай другие элементы ката БАССАЙ ДАИ удобнее всего смотреть на БЛОГГЕР-Константин Дробышевский-Бассай Дай Бункай, где представлены все варианты бункай: http://bassaidaibunkai.