CogniFit — Нейронная пластичность и когнитивность
«Под пластичностью мозга подразумевается способность нервной системы изменять свою структуру и функции на протяжении всей жизни в ответ на многообразие окружающей среды. Этому термину не так просто дать определение даже несмотря на то, что в настоящее время он широко применяется в психологии и нейронауке. Он используется для обозначения изменений, происходящих на различных уровнях нервной системы: в молекулярных структурах, изменения экспрессии генов и поведения».
Нейропластичность позволяет нейронам восстанавливаться как анатомически, так и функционально, а также создавать новые синаптические связи. Нейронная пластичность — это способность мозга к восстановлению и реструктуризации. Этот адаптивный потенциал нервной системы позволяет мозгу восстановиться после травм и нарушений, а также может уменьшить последствия структурных изменений, вызванных такими патологиями, как рассеянный склероз, болезнь Паркинсона, когнитивное расстройство, болезнь Альцгеймера, дислексия, СДВГ, бессонница у взрослых, бессонница у детей и т.
Нейронные сети до тренировокНейронные сети после 2-х недель когнитивной стимуляцииНейронные сети после 2-х месяцев когнитивной стимуляции
Синаптическая пластичность
Когда мы учимся или получаем новый опыт, мозг устанавливает серию нейронных связей. Эти нейронные сети представляют собой пути, по которым нейроны обмениваются между собой информацией. Эти пути формируются в мозге при обучении и практике, как, например, в горах образуется тропа, если по ней ежедневно ходит пастух со своим стадом. Нейроны взаимодействуют между собой посредством соединений, называемых синапсом, и эти коммуникационные пути могут восстанавливаться в течение всей жизни. Каждый раз, когда мы приобретаем новые знания (путем постоянной практики), коммуникация или синаптическая трансмиссия между участвующими в процессе нейронами усиливается. Улучшение коммуникации между нейронами означает, что электрические сигналы более эффективно передаются на протяжении всего нового пути. Например, когда вы пытаетесь распознать что за птица поёт, между некоторыми нейронами образуются новые связи. Так, нейроны зрительной коры определяют цвет птицы, слуховой коры — её пение, а другие нейроны — название птицы. Таким образом, чтобы идентифицировать птицу, нужно многократно сопоставить её цвет, голос, название. С каждой новой попыткой, при возвращении к нейронной цепи и восстановлении нейронной передачи между вовлечёнными в процесс нейронами, эффективность синаптической трансмиссии повышается. Таким образом, коммуникация между соответствующими нейронами улучшается, и процесс познания с каждым разом происходит быстрее. Синаптическая пластичность является основой пластичности человеческого мозга.
Например, когда вы пытаетесь распознать что за птица поёт, между некоторыми нейронами образуются новые связи. Так, нейроны зрительной коры определяют цвет птицы, слуховой коры — её пение, а другие нейроны — название птицы. Таким образом, чтобы идентифицировать птицу, нужно многократно сопоставить её цвет, голос, название. С каждой новой попыткой, при возвращении к нейронной цепи и восстановлении нейронной передачи между вовлечёнными в процесс нейронами, эффективность синаптической трансмиссии повышается. Таким образом, коммуникация между соответствующими нейронами улучшается, и процесс познания с каждым разом происходит быстрее. Синаптическая пластичность является основой пластичности человеческого мозга.
С учётом того, что синаптическая пластичность достигается путём улучшения коммуникаций в синапсе между существующими нейронами, под нейрогенезом подразумевается рождение и размножение новых нейронов в мозге. В течение длительного времени идея о регенерации нейронов в мозге взрослого человека считалась чуть ли не ересью. Учёные верили, что нервные клетки умирают и не восстанавливаются. После 1944 г., и особенно в последние годы, научным путём было доказано существование нейрогенеза, и сегодня мы знаем, что происходит, когда стволовые клетки (особый вид клеток, расположенных в зубчатой извилине, гиппокампе и, возможно, в префронтальной коре) делятся на две клетки: стволовую и клетку, которая превратится в полноценный нейрон, с аксонами и дендритами. После этого новые нейроны мигрируют в различные области (включая удалённые друг от друга) мозга, туда, где они нужны, поддерживая тем самым нейронную дееспособность мозга. Известно, что как у животных, так и у людей внезапная гибель нейронов (например, после кровоизлияния) является мощным стимулом для запуска процесса нейрогенеза.
Учёные верили, что нервные клетки умирают и не восстанавливаются. После 1944 г., и особенно в последние годы, научным путём было доказано существование нейрогенеза, и сегодня мы знаем, что происходит, когда стволовые клетки (особый вид клеток, расположенных в зубчатой извилине, гиппокампе и, возможно, в префронтальной коре) делятся на две клетки: стволовую и клетку, которая превратится в полноценный нейрон, с аксонами и дендритами. После этого новые нейроны мигрируют в различные области (включая удалённые друг от друга) мозга, туда, где они нужны, поддерживая тем самым нейронную дееспособность мозга. Известно, что как у животных, так и у людей внезапная гибель нейронов (например, после кровоизлияния) является мощным стимулом для запуска процесса нейрогенеза.
Функциональная компенсационная пластичность
В научной литературе по нейробиологии широко раскрыта тема снижения когнитивных способностей при старении и объяснено, почему пожилые люди демонстрируют более низкую когнитивную производительность, чем молодёжь. Удивительно, однако далеко не все пожилые люди показывают низкую производительность: у некоторых результаты ничуть не хуже, чем у молодых. Эти неожиданно разные результаты у подгруппы людей одного и того же возраста были исследованы научным путём, в результате чего было обнаружено, что при обработке новой информации пожилые люди с большей когнитивной производительностью используют те же самые области мозга, что и молодёжь, а также другие области мозга, которые не используются ни молодыми, ни другими пожилыми участниками эксперимента. Этот феномен сверхиспользования мозга пожилыми людьми был исследован учёными, которые пришли к выводу о том, что использование новых когнитивных ресурсов происходит в рамках компенсационной стратегии. В результате старения и снижения синаптической пластичности мозг, демонстрируя свою пластичность, начинает реструктурировать свои нейрокогнитивные сети. Исследования показали, что мозг приходит к этому функциональному решению путём активации других нервных путей, чаще задействуя области в обоих полушариях (что обычно характерно только для более молодых людей).
Удивительно, однако далеко не все пожилые люди показывают низкую производительность: у некоторых результаты ничуть не хуже, чем у молодых. Эти неожиданно разные результаты у подгруппы людей одного и того же возраста были исследованы научным путём, в результате чего было обнаружено, что при обработке новой информации пожилые люди с большей когнитивной производительностью используют те же самые области мозга, что и молодёжь, а также другие области мозга, которые не используются ни молодыми, ни другими пожилыми участниками эксперимента. Этот феномен сверхиспользования мозга пожилыми людьми был исследован учёными, которые пришли к выводу о том, что использование новых когнитивных ресурсов происходит в рамках компенсационной стратегии. В результате старения и снижения синаптической пластичности мозг, демонстрируя свою пластичность, начинает реструктурировать свои нейрокогнитивные сети. Исследования показали, что мозг приходит к этому функциональному решению путём активации других нервных путей, чаще задействуя области в обоих полушариях (что обычно характерно только для более молодых людей).
Функционирование и поведение: обучение, опыт и окружение
Мы рассмотрели, что пластичность — это способность мозга изменять свои биологические, химические и физические характеристики. Однако меняется не только мозг — также меняется поведение и функционирование всего организма. За последние годы мы узнали о том, что генетические или синаптические нарушения мозга происходят в результате как старения, так и воздействия огромного количества факторов окружающей среды. Особенно важны открытия о пластичности мозга, а также о его уязвимости в результате различных расстройств. Мозг учится на протяжении всей нашей жизни — в любой момент и по разным причинам мы получаем новые знания. Например, дети приобретают новые знания в огромных количествах, что провоцирует значительные изменения в мозговых структурах в моменты интенсивного обучения. Новые знания можно получить и в результате пережитой неврологической травмы, например, в результате повреждения или кровоизлияния, когда функции повреждённой части мозга нарушаются, и нужно учиться заново. Есть также люди с жаждой знаний, для которых необходимо постоянно учиться. В связи с огромным количеством обстоятельств, при которых может потребоваться новое обучение, мы задаемся вопросом, меняется ли каждый раз при этом мозг? Исследователи полагают, что это не так. По-видимому, мозг приобретает новые знания и демонстрирует свой потенциал пластичности в том случае, если новые знания помогут улучшить поведение. То есть для физиологических изменений мозга необходимо, чтобы следствием обучения были перемены в поведении. Другими словами, новые знания должны быть нужными. Например, знания о еще одном способе выживания. Вероятно, тут играет роль степень полезности. В частности, развить пластичность мозга помогают интерактивные игры. Было доказано, что такая форма обучения повышает активность префронтальной коры головного мозга (ПФК). Кроме того, полезно играть с положительным подкреплением и вознаграждением, что традиционно используется при обучении детей.
Есть также люди с жаждой знаний, для которых необходимо постоянно учиться. В связи с огромным количеством обстоятельств, при которых может потребоваться новое обучение, мы задаемся вопросом, меняется ли каждый раз при этом мозг? Исследователи полагают, что это не так. По-видимому, мозг приобретает новые знания и демонстрирует свой потенциал пластичности в том случае, если новые знания помогут улучшить поведение. То есть для физиологических изменений мозга необходимо, чтобы следствием обучения были перемены в поведении. Другими словами, новые знания должны быть нужными. Например, знания о еще одном способе выживания. Вероятно, тут играет роль степень полезности. В частности, развить пластичность мозга помогают интерактивные игры. Было доказано, что такая форма обучения повышает активность префронтальной коры головного мозга (ПФК). Кроме того, полезно играть с положительным подкреплением и вознаграждением, что традиционно используется при обучении детей.
Условия реализации пластичности мозга
Когда, в какой момент жизни мозг наиболее подвержен изменениям под воздействием факторов окружающей среды? По-видимому, пластичность мозга зависит от возраста, и предстоит сделать ещё немало открытий о влиянии на неё окружающей среды в зависимости от возраста субъекта.

1] Определение взято из: Колб, Б., Мохамед, A., & Гибб, Р., Поиск факторов, лежащих в основе пластичности мозга в нормальном и повреждённом состоянии, Revista de Trastornos de la Comunicación (2010), doi: 10. 1016/j.jcomdis.2011.04 0.007 Этот раздел является производным от работы Колба, B., Мохамеда, A., & Гибба, Р., Поиск факторов, лежащих в основе пластичности мозга в нормальном и повреждённом состоянии, Revista de Trastornos de la Comunicación (2010), doi: 10.1016 / j . jcomdis.2011.04.007
1016/j.jcomdis.2011.04 0.007 Этот раздел является производным от работы Колба, B., Мохамеда, A., & Гибба, Р., Поиск факторов, лежащих в основе пластичности мозга в нормальном и повреждённом состоянии, Revista de Trastornos de la Comunicación (2010), doi: 10.1016 / j . jcomdis.2011.04.007
Нейропластичность: перестраиваем мозг / Хабр
Наш мозг необычайно пластичен. Не как пластиковая посуда или кукла Барби – в неврологии пластичность означает удивительную способность мозга меняться и адаптироваться практически ко всему, что с нами происходит. В былые времена учёные считали, что когда человек переставал быть ребёнком, его мозг застывал, как глиняный горшок, и оставался в одной форме. Но кипы исследований опровергли их мнение – мозг больше напоминает пластилин [play-doh]. Эти изменения могут происходить на разных масштабах: от отдельного нейрона, меняющего связи, до целой корковой области, уменьшающейся или разбухающей. Структуру мозга могут менять множество факторов, от травм и инсультов, до медитации, упражнений или ежедневных занятий на пианино.
Начнём с небольших масштабов и синаптической пластичности (если вы не в курсе, что есть синапс, прочтите сначала вводную статью про мозг). Эта разновидность пластичности, которую часто называют длительной потенциацией (ДПЦ) и длительным подавлением (ДПД), критична для нашего понимания процессов запоминания и обучения. Очень упрощённо она работает так: связи между нейронами усиливаются или ослабляются (происходит потенциация или депрессия) в зависимости от их действий. Когда нейрон А постоянно возбуждает нейрон Б, связь между ними усиливается.
Естественно, обычно это происходит на нескольких синапсах – так могут появиться целые сети, в случае, если они достаточное количество раз именно в таком составе проявили активность (и мы считаем, что память формируется примерно так же). Так что целуйте вашу половинку достаточно часто во время прослушивания композиций Лу Бега, и скоро песня «Mambo number five» будет вызывать у вас романтические чувства. Дональд Хебб [Donald Hebb], канадский невропсихолог, придумал поговорку «Вместе запускаются, вместе заплетаются» [What fires together, wires together] для описания этого процесса. Изначально эти связи хрупкие, но если вы активируете их достаточное количество раз, они превратятся в прочные (их нельзя будет разделить, как Бритни и Джастина в 99-м). Обратный процесс, ДПД, запускается другой процедурой стимуляции, и, как считается, ослабляет ненужные связи – вы забываете имя вашей бывшей или облагораживаете новые танцевальные движения. Пластичность синапсов – концепция, которую когнитивные и поведенческие терапевты рекомендуют своим пациентам: для смены устоявшихся мысленных шаблонов нужно шаг за шагом при помощи практики формировать новые. И новые пути проходят эволюцию от грунтовых дорог до скоростных шоссе (по которым передвигается здоровое поведение), а сломанные контуры уплывают в небытие.
Так что целуйте вашу половинку достаточно часто во время прослушивания композиций Лу Бега, и скоро песня «Mambo number five» будет вызывать у вас романтические чувства. Дональд Хебб [Donald Hebb], канадский невропсихолог, придумал поговорку «Вместе запускаются, вместе заплетаются» [What fires together, wires together] для описания этого процесса. Изначально эти связи хрупкие, но если вы активируете их достаточное количество раз, они превратятся в прочные (их нельзя будет разделить, как Бритни и Джастина в 99-м). Обратный процесс, ДПД, запускается другой процедурой стимуляции, и, как считается, ослабляет ненужные связи – вы забываете имя вашей бывшей или облагораживаете новые танцевальные движения. Пластичность синапсов – концепция, которую когнитивные и поведенческие терапевты рекомендуют своим пациентам: для смены устоявшихся мысленных шаблонов нужно шаг за шагом при помощи практики формировать новые. И новые пути проходят эволюцию от грунтовых дорог до скоростных шоссе (по которым передвигается здоровое поведение), а сломанные контуры уплывают в небытие.
Пластичность на больших масштабах проявляется по-другому. Растущее количество исследований показывают, что чем больше вы используете определённый мускул, тем большую область мозг для него выделяет. Например, одно из исследований показывает, что хотя области, отвечающие за движения пальцев, обычно имеют одинаковый размер, это не неизменно. После пяти дней упражнений на пианино были найдены определённые и вполне видимые изменения в двигательной области коры мозга. Области, отвечающие за движения пальцев, расширились и заняли другие части соседних областей, будто сорняки, разрастающиеся в саду. Исследователи пошли ещё дальше: они показали, что даже если испытуемые думали об упражнениях, эффект получался почти таким же! Мысленные упражнения оказались настолько же эффективными в реорганизации структуры мозга, как и физические. Ещё один пример (о котором студенты-неврологи слышали, наверное, чаще, чем жители «Bible Belt» – областей США, где особенно силён протестантский фундаментализм – об Иисусе) – это лондонские таксисты. Опытные таксисты, которым приходится запоминать карту столицы, включая десятки тысяч улиц и десятки достопримечательностей, обладают большим задним гиппокампом – структурой мозга, отвечающей за пространственную память и ориентацией. У контрольной группы, водителей автобусов с постоянными и устоявшимися маршрутами, гиппокамп обычного размера. Чтобы предотвратить обычные комментарии типа «корреляция не гарантирует причинности» (ведь, возможно, именно увеличенный размер гиппокампа и привёл таксистов на эту работу?), исследователи показали, что увеличении объёма гиппокампа позитивно коррелировала со временем, проведённым за баранкой. Чем дольше вы водите, тем больше ваш мозг адаптируется.
Опытные таксисты, которым приходится запоминать карту столицы, включая десятки тысяч улиц и десятки достопримечательностей, обладают большим задним гиппокампом – структурой мозга, отвечающей за пространственную память и ориентацией. У контрольной группы, водителей автобусов с постоянными и устоявшимися маршрутами, гиппокамп обычного размера. Чтобы предотвратить обычные комментарии типа «корреляция не гарантирует причинности» (ведь, возможно, именно увеличенный размер гиппокампа и привёл таксистов на эту работу?), исследователи показали, что увеличении объёма гиппокампа позитивно коррелировала со временем, проведённым за баранкой. Чем дольше вы водите, тем больше ваш мозг адаптируется.
Вы уже согласны с тем, что мозг невероятно пластичен? Не спешите, у нас есть ещё примеры. Если вы отказались от медитации, как от хипповой ерунды, обратите внимание: долговременная практика медитации связана с весьма положительными изменениями в мозгу. Воспринимайте её как тренировку – как уроки на пианино. Исследования показывают, что если сидеть спокойно и медитировать, то можно увеличить толщину коры (то есть, больше серых клеточек, то есть, больше нейронов для обработки сигналов) в областях, связанных с вниманием, памятью и управлением эмоциями. Более того, миндалина, центр реакций, связанных со страхом и отвращением, уменьшается и ослабляет связи с префронтальной корой головного мозга, местом, где расположены высшие исполнительные функции. Проще говоря, медитация позволяет реагировать на стресс более вдумчиво и подавлять инстинкты. Последнее по очереди, но не по значимости – сеть пассивного режима работы мозга, ответственная за самоопределение и грёзы наяву, также снижает активность, что позволяет меньше отвлекаться (и предотвращает перескакивание мыслей со вчерашней вечеринки на неотвратимость смерти или что-то подобное). И пока я тут занимаюсь скрытой пропагандой ЗОЖ, упомяну, что ещё меняют ваш мозг к лучшему физические упражнения. Всего три часа быстрой ходьбы в неделю увеличивает рост и рождение нервных клеток, что, в свою очередь, предотвращает возрастное уменьшение мозга.
 Исследование показывают, что особенно выигрывают от этого передние области и гиппокамп – то есть, их объём после длительных упражнений вырастал. Вот вам и пример того, как память и способность рассуждать улучшаются благодаря ЗОЖ.
Исследование показывают, что особенно выигрывают от этого передние области и гиппокамп – то есть, их объём после длительных упражнений вырастал. Вот вам и пример того, как память и способность рассуждать улучшаются благодаря ЗОЖ.
Ваш мозг, словно идеальный супруг, существует с вами вместе в хорошие времена и в плохие, в болезни и в здоровье. После перенесённой травмы или инсульта нейропластичность помогает вам. Реабилитационные тренировки после инсульта или травмы показали, что мозг проводит реорганизацию вокруг повреждённого региона. Допустим, инсульт повредил часть мозга, отвечающую за движения левой руки. Использование технологии под названием «терапия принудительных двигательных ограничений» (когда вас заставляют пользоваться «плохой» рукой, в то время как другую руку ограничивают в движениях), ведёт к увеличению объёма серого вещества в двигательном отделе, меняет смежные с повреждённым регионы так, что они берут на себя его функции и даже заставляют контралатеральную полусферу участвовать в восстановлении. Мозг перестраивает себя, чтобы приспособиться к новым обстоятельствам и сделать это наилучшим образом. Однако, не всегда это проходит так здорово. Иногда мозг может подложить свинью и доставить вам неприятностей – это я насчёт фантомных болей. Вы, наверно, слышали о людях, у которых сохраняется ощущение ампутированных рук или ног. Это также заслуга нашего неугомонного пластичного мозга, хотя на 100% этот процесс не изучен. Одна из общепринятых теорий говорит, что область соматосенсорной коры, соседствующая с той, что отвечала за функции отсутствующей конечности, хватается за новую возможность и занимает вакантное место. К примеру, область лица расположена рядом с областью рук. И если потерять руку, область лица занимает место своего соседа и воспринимает все ощущения лица вдвойне: как идущие от щеки, так и от несуществующего большого пальца.
Мозг перестраивает себя, чтобы приспособиться к новым обстоятельствам и сделать это наилучшим образом. Однако, не всегда это проходит так здорово. Иногда мозг может подложить свинью и доставить вам неприятностей – это я насчёт фантомных болей. Вы, наверно, слышали о людях, у которых сохраняется ощущение ампутированных рук или ног. Это также заслуга нашего неугомонного пластичного мозга, хотя на 100% этот процесс не изучен. Одна из общепринятых теорий говорит, что область соматосенсорной коры, соседствующая с той, что отвечала за функции отсутствующей конечности, хватается за новую возможность и занимает вакантное место. К примеру, область лица расположена рядом с областью рук. И если потерять руку, область лица занимает место своего соседа и воспринимает все ощущения лица вдвойне: как идущие от щеки, так и от несуществующего большого пальца.
Становится понятно, что мы не ограничены теми картами, что нам раздала природа: возможно поменять некоторые из них (и это даже не будет воспринято, как шулерство). Мозг отражает наше окружение, наши решения, эмоции и стиль жизни, и поменять всё это, на самом деле, никогда не поздно.
Мозг отражает наше окружение, наши решения, эмоции и стиль жизни, и поменять всё это, на самом деле, никогда не поздно.
Источники:
LTP
Piano and plasticity 1, 2
Taxi drivers
Meditation and DMN
Meditation and amygdala
Meditation and increased cortical thickness
Brain and exercise 1, 2
Constraint-induced movement therapy
Phantom pain
A great meditation resource suited for beginners which I’m using
Pictures by Toma
Интересные моменты из «Пластичности мозга» Нормана Дойджа
Книга вызвала смешанные чувства. Плюсы: много новых идей, 2007 год, автор не журналист, а учёный, много ссылок на исследования. Минусы: повествование скачет с темы не тему, местами есть сомнительные утверждения. Чтобы во всём лучше разобраться и запомнить, я сначала составил карту, а потом и этот пост. Поехали.
Раньше считали, что мозг не способен меняться
Или способен, но только в сторону ухудшения. Считали, что со временем нейроны теряют способность правильно развиваться, получают повреждения или умирают.
Считали, что со временем нейроны теряют способность правильно развиваться, получают повреждения или умирают.
Оказалось, что ещё как способен
Возможно восстановление зрения через тактильные ощущения, когда камера транслирует изображение в вибрирующие с разной силой и частотой ощущения на спине, а человек учится видеть пространственное изображение с перспективой, читать, распознавать предметы и людей.
Восстановление слуха у полностью глухих людей — почти обычное дело. Полностью искусственный имплант подаёт на вход электрические сигналы, а мозг вполне неплохо распознаёт их как звук.
Эксперименты показывают, что мозг может научиться воспринимать любой поток сенсорной информации. И не только сенсорной. Мозг может быть сильно повреждён в результате травмы или инсульта, а пациент восстанавливает одну функцию за другой — учится управлять руками и ногами, ползать, ходить, говорить и писать.
Как исследуют мозг
Самый точный инструмент исследования — микроэлектроды, с помощью которых активируют отдельные нейроны или следят за их активацией и составляют «карты мозга» — какие нейроны за что отвечают.
С помощью картирования выяснили, что участки лобных долей мозга отвечают за двигательную систему. Височная, теменная и затылочная доли образуют сенсорную систему, которая обрабатывает сигналы всех огранов чувств. На карте мозга также отражается психическая активность более высокого уровня: например, память и воображение.
Принципы нейропластичности и пластический парадокс
Карты мозга меняются постоянно. Не используемая область мозга захватывается соседними и начинает обрабатывать информацию от них. Если мы прекращаем тренировать одни ментальные навыки, то пространство карты мозга, предназначенной для этих навыков, переходит к тем навыкам, которые мы продолжаем использовать.
Плохие привычки получают преимущество: каждое исполнение усиливает её и вытесняет смежные, а чтобы избавиться, надо долго не «кормить», то есть не следовать ей.
Принцип нейропластичности:
- — то, что мы используем, разрастается,
- — то, что мы не используем, сокращается.

Метафора снежной горы и санок
Нейропластичность похожа на мягкий снег, покрывающий гору.
Съезжая с горы на санках, мы можем быть гибкими, выбирая любой путь в мягком снегу. Однако когда мы используем одну и ту же проторенную трассу во второй или третий раз, мы оказываемся заложниками её колеи. Теперь наш маршрут становится неизменным, так как сложившиеся нейронные сети стремятся стать самодостаточными.
Чем чаще мозг пользуется каким-то маршрутом для спуска с горы, тем более он быстрый и эффективный, но тем сложнее его изменить.
Так формируются и застывают привычки: и хорошие, и плохие.
Это пластический парадокс
Пластичность мозга делает нас более гибкими, и она же делает нас менее гибкими.
Мозг можно тренировать так же, как мышцы
При этом мозг растёт физически! Например, тренировки мозга у крыс повышают объём мозга на 5%, кол-во нейронов на 9% и кол-во связей между нейронами на 25%.
Существуют фаза обучения и фаза закрепления
Сначала количество активируемых нейронов растёт — это фаза обучения. Например, при игре на фортепиано у пианиста активируются области пальцев, кисти, руки и плеча.
Например, при игре на фортепиано у пианиста активируются области пальцев, кисти, руки и плеча.
Потом количество задействованных нейронов сокращается — это фаза закрепления. Хотя очевидного прогресса на этапе закрепления не видно, в этот период происходят внутренние биологические изменения, в ходе которых навыки совершенствуются и приобретают автоматический характер. Пианист учится нажимать на клавиши одними пальцами. Точность растёт. Скорость растёт.
Есть краткосрочные и долгосрочные изменения от обучения
Учёные провели такой эксперимент: дали слепым людям учить азбуку Брайля по несколько занятий в неделю в течение года и измеряли задействованные при чтении зоны мозга.
К пятницам задействованные при чтении зоны мозга растут. Но к понедельникам возвращаются обратно. Это «эффект зубрёжки» — легко выучить, легко забыть.
«Понедельничные» зоны вырастали только через 6 месяцев. И сохранялись даже после двухмесячного перерыва в занятиях. Это долгосрочные изменения.
Эффективный способ обучения — барьеры
Например, здоровый человек с завязанными глазами буквально за 2 дня учится ориентироваться в пространстве с помощью тактильных ощущений и слуха. А при реалибитации инсульта завязанная здоровая рука помогает быстро разработать больную.
То же применимо к нашей работе: делать больше задач за неделю, уложиться к более жёсткий дедлайн, добавить ограничения. Ограничения рулят.
Обучение работает лучше, когда оно происходит чаще, дольше и интенсивнее
Лучше каждый день подряд, чем столько же дней с перерывами.
Лучше один час очень интенсивных занятий, чем 6 часов менее интенсивных.
Оптимально заниматься 3 часа в день.
Важную роль играет внимание
Многочисленные эксперименты показали, что изменения в картах активируемых нейронов происходили только тогда, когда испытуемые проявляли неослабный интерес к происходящему. Когда испытуемые выполняли задания автоматически, не уделяя этому особого внимания, карты их мозга менялись, но эти изменения длились недолго.
Чем бы вы ни занимались, должно быть интересно! Иначе ничего не получится. Как делать интересно — отдельная тема.
Важно идти от простого к сложному
Слишком просто — не интересно. Слишком сложно — не интересно. А когда не интересно, обучение не происходит.
Воображаемые тренировки могут быть так же эффективны, как и физические
Провели эксперимент. Взял две группы людей, которые никогда не учились игре на пианино, и показали им, как играть определённую последовательность нот, какими пальцами нажимать клавиши и как они звучат.
Дальше первая группа занялась «психической тренировкой»: пять дней по два часа они сидели перед пианино и представляли, как играют выученную последовательность нот и как слышат своё исполнение. А вторая группа занялась «физической тренировкой»: пять дней по два часа в день они действительно играли эту мелодию на пианино.
Затем представителей обеих групп попросили сыграть предложенную последовательность нот, а компьютер оценивал точность исполнения.
В группе с психическими тренировками, прогресс за пять дней был существенным, но менее значительным, чем у группы с физическими тренировками. Однако после того как первая группа закончила психические тренировки, и для ее участников был проведен двухчасовой сеанс физической тренировки, общий уровень исполнения участников обеих групп сравнялся
С точки зрения нейрофизиологии, мысленное представление какого-либо действия и его совершение не очень отличаются. Когда люди закрывают глаза и представляют простой объект, такой как буква «а», активируется первичная зрительная кора, словно они действительно смотрят на эту букву. Сканирование мозга показывает, что в процессе действия и его мысленного представления происходит активация ряда одинаковых участков мозга. Именно поэтому визуализация может улучшить исполнение.
Примеры из жизни
Вспоминание ответов на вопросы теста перед экзаменом, заучивание текста роли, репетиция выступления — всё это психические тренировки.
Этим можно и нужно пользоваться и в работе. Мысленные репетиции переговоров помогают прокачать навыки реальных переговоров. Проработка сценариев развития проекта учит управлять реальными проектами. Упражнение «взгляд новичка» на рекламу и сайт помогает лучше понимать пользователей.
Про старение и слабоумие
Вероятность болезни Альцгеймера (слабоумия) к 85 годам — 50%. И там ещё целый веер подобных заболеваний. Ведут к ухудшению памяти, мышления и скорости обработки информации.
Основная проблема — снижение скорости обработки информации. Из-за этого мы не успеваем фиксировать события → снижается точность и острота восприятия → невозможно точно запомнить → всё в голове становится «смутным» → и со временем всё восприятие, мысли, эмоции, решения, действия становятся всё менее точными.
Что делать?
1. Ни в коем случае не допускать возрастания «мутности» в голове
Если не успеваешь — останавливаться и разбираться, пока не разберёшься до конца. Не усиливать мутность алкоголем, недосыпом, хроническим стрессом и однообразием.
Не усиливать мутность алкоголем, недосыпом, хроническим стрессом и однообразием.
2. Тренировать голову
Подходят занятия, требующую высокой концентрации внимания: обучение игре на музыкальном инструменте, настольные игры, вдумчивое чтение, танцы.
Боулинг, присмотр за детьми и гольф не подходят.
3. Тренировать тело
Важна ходьба. Эксприменты на мышах показали, что количество новых нейронов увеличивается, если много ходить. Другая физическая активность мышей такого эффекта не даёт. Людям, вероятно, подойдёт также бег, езда на велосипеде, плавание.
Любые кардиоваскулярные упражнения также полезны, потому что дают мозгу больше кислорода, стимулируют выделение BDNF и помогают чувствовать себя в лучшей психической форме.
Но главное — учиться чему-то действительно новому.
Купить: электронная, аудио, на бумаге.
Пластичность мозга: ограничения и возможности
Пластичностью мозга (ПМ) принято называть динамичное структурно-функциональное реагирование мозга на изменения внешней среды и самого организма. Механизмы П.М. обеспечивают также реагирование на повреждения самого мозга и периферической нервной сети и восстановление утраченных функций [1, 2]. В числе наиболее частых форм патологии мозга — инсульт, поэтому далее ПМ будет рассматриваться в аспекте этого заболевания.
Механизмы П.М. обеспечивают также реагирование на повреждения самого мозга и периферической нервной сети и восстановление утраченных функций [1, 2]. В числе наиболее частых форм патологии мозга — инсульт, поэтому далее ПМ будет рассматриваться в аспекте этого заболевания.
Восстановительный потенциал ПМ при инсульте определяется прежде всего возрастом пациента, длительностью воздействия повреждающего агента, локализацией повреждения мозга, масштабами нарушения гематоэнцефалического барьера и балансом между развернувшейся на повреждение мозга реакцией иммунной системы (ИС) и механизмами нейропротекции. Однако механизмы ПМ могут быть и виновниками развития патологических состояний. С сетевой структурной организацией связано такое негативное проявление ПМ, как нейродегенерация в нем функционально связанных с зоной повреждения отдаленных интактных областей [3]. Обширность поражения мозга, индивидуальные вариации анатомических особенностей и функциональных связей мозга также относятся к числу факторов, определяющих П. М. Исследования последних лет по эпигенетической регуляции [4] расширили наши представления о молекулярных механизмах ПМ при физиологических и патологических процессах.
М. Исследования последних лет по эпигенетической регуляции [4] расширили наши представления о молекулярных механизмах ПМ при физиологических и патологических процессах.
Неонейрогенез происходит с различной интенсивностью на протяжении всей жизни млекопитающих в передней части субвентрикулярной зоны и в субгранулярной зоне зубчатой извилины гиппокампа. Существование клеток-предшественников в различных отделах коры и подкорковых образованиях не исключает возможности проявления ПМ в медленном ограниченном самообновлении мозга на протяжении жизни человека. Однако при наличии эндогенного потенциала неонейрогенеза и восстановления аксонов при большинстве заболеваний мозга эффективной регенерации его не происходит, что чаще всего объясняют доминированием реактивного глиоза при ответе на локальное воспаление, обусловленным, по-видимому, пространственно-временны́м диссонансом между участниками регенеративного процесса [5].
Рассмотрим перспективы использования трансплантации стволовых клеток в мозг и новые возможности влияния на механизмы ПМ и регенерации с учетом существующих ограничений.
Особенность развития коры мозга всех млекопитающих состоит в том, что ни один из составляющих ее нейронов не образуется в пределах самой коры, а мигрирует из пролиферативных транзиторных эмбриональных зон (в частности, из вентрикулярных и субвентрикулярных зон, расположенных вблизи боковых желудочков). Слоение коры происходит не только за счет миграции постмитотических нейронов, но и достраиванием ее слоев (за исключением слоя I) в последовательности изнутри—кнаружи. Биологическое значение этого феномена не ясно, но в разработке новых подходов к регенерации мозга следует иметь в виду, что нарушение последовательности изнутри—кнаружи ведет к нарушению кортикальной функции [6]. К тому же развитие функций мозга в постнатальном периоде связанно с возрастными пространственно-временны́ми ограничениями в нем, обусловленными особенностями разных его отделов и взаимосвязей между ними.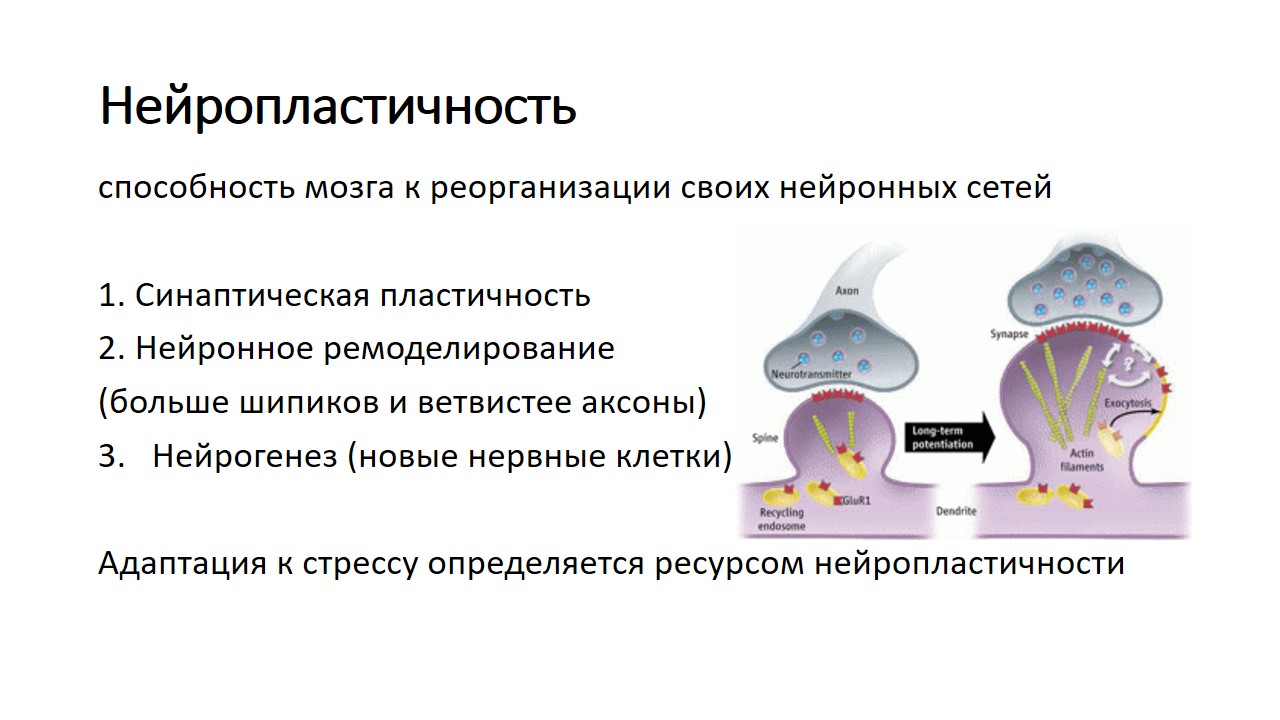 Несформировавшаяся в соответствующий момент раннего развития ребенка функция мозга (например речь) часто не развивается нормально либо формируется в более поздние сроки. Мозг оказывается уже на другом этапе развития и невозможно воспроизведение предшествовавших стадий структурно-функциональных отношений, соответствующих развитию и становлению этой функции.
Несформировавшаяся в соответствующий момент раннего развития ребенка функция мозга (например речь) часто не развивается нормально либо формируется в более поздние сроки. Мозг оказывается уже на другом этапе развития и невозможно воспроизведение предшествовавших стадий структурно-функциональных отношений, соответствующих развитию и становлению этой функции.
Первоначально выявление потенциальной способности мозга к самообновлению породило всплеск исследований по нейрональной заместительной терапии. Большинство работ в этом направлении были выполнены на животных и по многим причинам возможности переноса их результатов на человека оказались весьма ограниченными. Так излюбленным и наиболее экономичным подходом в изучении патологических процессов человеческого мозга чаще всего служит моделирование их на грызунах, что оправдывают большим сходством мозга млекопитающих, игнорируя при этом существование различий не только размеров самого мозга и специфичных только для человека областей коры, в частности областей, связанных с речью, но и источники, и пути миграции нейронов в определенные области мозга, которые невозможно смоделировать не только на грызунах, но и даже на приматах. Пренебрежение эволюционными различиями мозга, возможно, служит одним из объяснений неудач трансляций в клинику перспективных результатов, полученных на грызунах. При отличии по специфическим генам между человеком и грызунами в 1%, разница между ними по времени, последовательности и уровню экспрессии генов слишком велика [6].
Пренебрежение эволюционными различиями мозга, возможно, служит одним из объяснений неудач трансляций в клинику перспективных результатов, полученных на грызунах. При отличии по специфическим генам между человеком и грызунами в 1%, разница между ними по времени, последовательности и уровню экспрессии генов слишком велика [6].
Нельзя не упомянуть о других предостережениях. Как правило, в экспериментах используют более молодых животных. Им свойственна низкая коморбидность, и моделируют на них более гомогенные по характеру повреждения мозга. Чаще всего используются грызуны, которым свойственно передвижение на четырех конечностях, а ориентирование у них в значительной степени происходит с участием вибрисс. Нельзя игнорировать различия мозга человека и грызунов по объему белого вещества, как и размеры и структурную сложность мозга в целом [7, 8].
Острая воспалительная реакция клеток врожденной ИС мозга на его повреждение ингибирует образование и сохранение новых нейронов у взрослого. Эволюционное давление выбрало, по-видимому, в качестве реакции на повреждение ту, которая благоприятствует ремиелинизации и спасению существовавшей до повреждения нейронной сети мозга, а не образование в нем полностью новых сетей. Тонкая функциональная настройка сетей мозга и его созревание у человека занимают, по-видимому, два десятилетия. Экспериментально показано, что трансплантированные нейрональные стволовые клетки обладают способностью опосредовать ограниченное восстановление в ЦНС, проявляющееся главным образом образованием олигодендроцитов и астроцитов, и редко полным восстановлением функции поврежденного мозга. В аспекте отмеченных выше особенностей эмбриогенеза мозга млекопитающих и постнатального развития мозга человека и эволюционно закрепленной его реакции на повреждение наивно предполагать, что мозг будет эффективно использовать инъецированные стволовые клетки как субстраты для регенерации нейронных сетей. Пораженный мозг не способен эффективно возмещать утраченные нейроны без агрессивных интервенций, которые преодолевали бы естественное ингибирование в нем нейрогенеза [9].
Эволюционное давление выбрало, по-видимому, в качестве реакции на повреждение ту, которая благоприятствует ремиелинизации и спасению существовавшей до повреждения нейронной сети мозга, а не образование в нем полностью новых сетей. Тонкая функциональная настройка сетей мозга и его созревание у человека занимают, по-видимому, два десятилетия. Экспериментально показано, что трансплантированные нейрональные стволовые клетки обладают способностью опосредовать ограниченное восстановление в ЦНС, проявляющееся главным образом образованием олигодендроцитов и астроцитов, и редко полным восстановлением функции поврежденного мозга. В аспекте отмеченных выше особенностей эмбриогенеза мозга млекопитающих и постнатального развития мозга человека и эволюционно закрепленной его реакции на повреждение наивно предполагать, что мозг будет эффективно использовать инъецированные стволовые клетки как субстраты для регенерации нейронных сетей. Пораженный мозг не способен эффективно возмещать утраченные нейроны без агрессивных интервенций, которые преодолевали бы естественное ингибирование в нем нейрогенеза [9]. Проблема состоит в обеспечении полной анатомической и функциональной интеграции новых нейронов в пораженной области. Сложность интеграции определяется и тем, что введенные нейрональные стволовые клетки должны быть признаны как «свои», быть не только регион-специфичными, но и соответствовать слою коры и быть нейронами определенного типа, причем способными формировать длинные связи. По-видимому, из-за этих фундаментальных препятствий в стартовавших крупнейших европейском и американском проектах предусмoтрено изучение лишь индуцибельных плюрипотентных стволовых клеток [10, 12].
Проблема состоит в обеспечении полной анатомической и функциональной интеграции новых нейронов в пораженной области. Сложность интеграции определяется и тем, что введенные нейрональные стволовые клетки должны быть признаны как «свои», быть не только регион-специфичными, но и соответствовать слою коры и быть нейронами определенного типа, причем способными формировать длинные связи. По-видимому, из-за этих фундаментальных препятствий в стартовавших крупнейших европейском и американском проектах предусмoтрено изучение лишь индуцибельных плюрипотентных стволовых клеток [10, 12].
Понимание регенерационной способности мозга должно опираться, как отмечено нами ранее [1], на биологические предпосылки. Каждый орган поддерживает свой клеточный гомеостаз в том временно́м ритме, который согласован с его функциональными возрастными потребностями и запрограммированной стабильностью составляющих его клеточных элементов, внося свой вклад по обеспечению целостности организма. По рейтингу регенерационной способности мозгу обычно присваивают низшую оценку. Вместе с тем присутствие клеток-предшественников практически во всех областях мозга следует рассматривать не как излишество, а как свидетельство их обеспечения регенерационным потенциалом. Мозг наделен регенерационной способностью в той мере, в какой это целесообразно в эволюционном аспекте развития вида, и она подвержена изменениям. Присущая мозгу пластичность в числе различных механизмов активно использует, по-видимому, и регенерационные процессы, успешно устраняя возникающие в его структуре лишь малые повреждения, остающиеся для нас незамеченными.
Вместе с тем присутствие клеток-предшественников практически во всех областях мозга следует рассматривать не как излишество, а как свидетельство их обеспечения регенерационным потенциалом. Мозг наделен регенерационной способностью в той мере, в какой это целесообразно в эволюционном аспекте развития вида, и она подвержена изменениям. Присущая мозгу пластичность в числе различных механизмов активно использует, по-видимому, и регенерационные процессы, успешно устраняя возникающие в его структуре лишь малые повреждения, остающиеся для нас незамеченными.
Эволюционные ограничения в регенерационной способности мозга связаны с реализацией его функций. Учитывая широкий временно́й диапазон функциональных процессов мозга, ограниченный нейрогенез при развитии и во взрослом состоянии, по-видимому, связан не с быстрыми процессами, а с механизмами медленной ПМ, закрепляющей, например, овладение сложными профессиональными умениями. Трудно представить, как человек существовал бы в природе при быстром клеточном обновлении мозга. Невозможно было бы сохранять в памяти необходимые жизненные навыки, информацию об окружающем мире и ориентироваться в нем. Резко ограничены были бы комбинаторные механизмы мозга, лежащие в основе сознания, мышления, языка и базирующиеся на механизмах памяти. Неонейрогенез в гиппокампе, как свидетельствуют новые экспериментальные данные и компьютерное моделирование, может нарушать процессы памяти. С биологической точки зрения в мозге недопустим регенерационный механизм, действующий, например в печени либо при непрерывном обновлении элементов крови. Поэтому в ответах на такие повреждения мозга, как травма или инсульт, превалирует эволюционно сложившееся «предпочтение» в пользу ремиелинизации и сохранения (спасения) существовавшей нейронной сети. Воссоздание полностью новых сетей в очаге поражения аннулировало бы сформировавшуюся ранее под воздействием обучения и адаптации тонкую настройку сетей и привело бы к утрате многих функций, по крайней мере до того момента, пока новая сеть не «переобучится» и не интегрируется в сохранившиеся сети мозга.

Внимание исследователей ПМ не могла не привлечь эпигенетика, предметом которой является изучение митотических и мейотических наследуемых изменений в функционировании генома, которые не могут быть объяснены изменениями первичной структуры ДНК. Они обусловлены структурными адаптационными изменениями в хромосомах и фиксируют состояния их измененной активности, возникая на протяжении всей жизни — от эмбриональной стадии и до старости — как результат развития физиологических процессов, влияния окружения, питания или, что хотелось бы подчеркнуть, случайного события [4].
По своему проявлению они могут быть такими же сильными, как и генетические эффекты, обнаруживаются в многостадийной и многоуровневой дезорганизации организма, передаются нескольким поколениям по мужской и женской линиям, а при определенных обстоятельствах также реверсируются. В осуществлении их задействованы различные молекулярные процессы: метилирование нуклеиновых кислот, модификация гистонов, позиционирование нуклеосом на ДНК, транскрипционный контроль связывающимися с ДНК белками и некодирующими (нк) малыми и длинными РНК, трансляционный контроль микроРНК (~22 нуклеотида) и связывающимися с РНК белками. Среди них наиболее изученными являются метилирование ДНК, происходящее в цитозиновом основании в контексте дуплета CpG, и модификации гистонов (метилирование, ацетилирование, фосфорилирование и др.) [4, 12—15]. Особенно сложна посттранcкрипционная регуляция. Она представлена сложной сетью взаимодействий между различными типами РНК: белок-кодирующими матричными РНК и нкРНК (длинными нкРНК, псевдогенами и циркулярными РНК). Эти РНК-транскрипты выступают как конкурирующие эндогенные «губки» микроРНК: они сообщаются и регулируют друг друга, конкурируя за связывание комплементарных к ним микроРНК [16]. Открытые первоначально в клетках как регуляторы экспрессии генома и трансляции белков, микроРНК циркулируют в стабильной форме в разных жидкостях организма и могут служить в качестве нового поколения диагностических и прогностических биомаркеров. Новые классы малых и длинных нкРНК характеризуются участием в широком спектре физиологических и патологических процессов.
Среди них наиболее изученными являются метилирование ДНК, происходящее в цитозиновом основании в контексте дуплета CpG, и модификации гистонов (метилирование, ацетилирование, фосфорилирование и др.) [4, 12—15]. Особенно сложна посттранcкрипционная регуляция. Она представлена сложной сетью взаимодействий между различными типами РНК: белок-кодирующими матричными РНК и нкРНК (длинными нкРНК, псевдогенами и циркулярными РНК). Эти РНК-транскрипты выступают как конкурирующие эндогенные «губки» микроРНК: они сообщаются и регулируют друг друга, конкурируя за связывание комплементарных к ним микроРНК [16]. Открытые первоначально в клетках как регуляторы экспрессии генома и трансляции белков, микроРНК циркулируют в стабильной форме в разных жидкостях организма и могут служить в качестве нового поколения диагностических и прогностических биомаркеров. Новые классы малых и длинных нкРНК характеризуются участием в широком спектре физиологических и патологических процессов.
В нервной системе эпигенетическая регуляция экспрессии генов играет роль в развитии и дифференциации мозга, синаптической пластичности, обучении и памяти, нейропротекции, в патологических проявлениях, связанных с нарушениями развития нервной системы (синдром Ретта, синдром хрупкой Х-хромосомы и др. ), психическими (наркомания, шизофрения, депрессия и др.) и нейродегенеративными (болезнь Паркинсона, болезнь Альцгеймера и др.) заболеваниями, опухолевым ростом и старением. В моделях ишемии мозга и инсульта препараты, влияющие на различные уровни ЭР (например, ингибиторы гистоновых деацетилаз), проявляли нейропротективный эффект. При окклюзии мозговых артерий, как модели ишемии, ацетилирование гистона Н3 было выраженно сниженным [4].
), психическими (наркомания, шизофрения, депрессия и др.) и нейродегенеративными (болезнь Паркинсона, болезнь Альцгеймера и др.) заболеваниями, опухолевым ростом и старением. В моделях ишемии мозга и инсульта препараты, влияющие на различные уровни ЭР (например, ингибиторы гистоновых деацетилаз), проявляли нейропротективный эффект. При окклюзии мозговых артерий, как модели ишемии, ацетилирование гистона Н3 было выраженно сниженным [4].
В аспекте многочисленных механизмов эпигенетической регуляции на уровне транскрипции и трансляции, которые можно принять как нижний уровень регуляции, патофизиологические модели, например инсульта, отображают с разной степенью полноты лишь верхний уровень изменений, проявляющихся в нарушенной межклеточной коммуникации, гибели клеток в поврежденной области и объемной диффузии продуктов их распада, запускающих каскад патогенетических механизмов [17]. Влияние последних на нижний уровень регуляции может быть плейотропным и сложным, и лишь в последнее время он стал рассматриваться в качестве перспективной мишени терапевтического воздействия при инсульте. Возникающие при поражении мозга симптомы отражают не столько проявления повреждения его, сколько состояние всего мозга, а точнее пластические изменения в оставшейся функционирующей части мозга [1], в числе которых могут быть и те, которые блокируют восстановление функции. Используемой терапией не удается после инсульта остановить прогрессирование размеров зоны инфаркта. Возможно, это объясняется необратимостью изменений на уровне Э.Р. Кроме того, воздействие только на какую-то одну мишень может оказаться малоэффективным из-за дефекта патогенетических механизмов, т. е. из-за поддержания патологического процесса разными молекулярными системами, когда выключение одной из них не ослабляет и не блокирует его.
Возникающие при поражении мозга симптомы отражают не столько проявления повреждения его, сколько состояние всего мозга, а точнее пластические изменения в оставшейся функционирующей части мозга [1], в числе которых могут быть и те, которые блокируют восстановление функции. Используемой терапией не удается после инсульта остановить прогрессирование размеров зоны инфаркта. Возможно, это объясняется необратимостью изменений на уровне Э.Р. Кроме того, воздействие только на какую-то одну мишень может оказаться малоэффективным из-за дефекта патогенетических механизмов, т. е. из-за поддержания патологического процесса разными молекулярными системами, когда выключение одной из них не ослабляет и не блокирует его.
Сложность изучения вклада эпигенетической регуляции в ПМ заключается в том, что мозг — одна из наиболее гетерогенных структур организма, его различные области неодинаково реагируют на одни и те же внешние и внутренние изменения, прием лекарственных препаратов, в одной и той же области разные типы ее клеток по-разному реагируют на один и тот же фактор в зависимости от половых различий [2]. Кроме того, как говорилось выше, множественны сами механизмы эпигенетической регуляции.
Кроме того, как говорилось выше, множественны сами механизмы эпигенетической регуляции.
При ограниченной регенерационной способности мозга важным представляется более полное использование механизмов ПМ, реализующих восстановление его поврежденных функций. В дополнение к традиционно используемой нейрофармакотерапии в последние годы показано успешное использование физического и когнитивного тренинга, глубокой стимуляции мозга, факторов роста при поражениях мозга. Если ранее точка зрения на патогенез многих поражений мозга, как и на процессы, связанные с восстановлением мозга, имела преимущественно «нейроцентрическую» направленность, сегодня же признано активное участия в них сосудистой системы и И.С. Одним из свидетельств этому служит, к примеру, то, что увеличение уровня эндотелиальных прогениторных клеток после ишемического инсульта коррелировало с хорошим функциональным исходом и уменьшенным образованием зоны инфаркта мозга [18].
Из физиологической интегрированности нервной, иммунной и сосудистой систем очевидно, что проблема восстановления мозга должна решаться интегративно, охватывая каждую систему. Наиболее перспективны в этом аспекте воздействия на ПМ неинвазивной транскраниальной магнитной стимуляцией (ТМС) и транскраниальной микроэлектростимуляцией (ТМЭС) мозга [19—22]. Последнее пятилетие характерно всплеском публикаций по успешному использованию их не только при инсульте для стимуляции эндогенных механизмов регенерации. Оба типа воздействия позволяют избирательно стимулировать мозг в различных режимах, активируя разные функции: моторику, память, речь. Под контролем нейровизуализации можно проследить изменения в картине активации крупномасштабных сетей мозга и целенаправленно корректировать терапевтический эффект.
При сравнении достоинств обоих методов следует особо упомянуть ограничения, связанные с использованием ТМС. Ее непосредственное воздействие приходится лишь на поверхностные слои мозга и слабо затрагивает глубокие отделы белого вещества. Невозможно обеспечить посредством ТМС одновременное воздействие на разные области мозга крупномасштабной сети, восстановить активность в длиннодистантных ассоциативных трактов белого вещества (например, тракты, соединяющие фронтальные и затылочные либо фронтальные и теменные области мозга), воздействуя на них одновременно с разных концов в пределах полушария, либо в комиссуральных трактах, воздействуя одновременно на оба полушария. Кроме того, у ТМС меньшие возможности варьирования параметров магнитного поля и локализации стимуляции. Повторные сеансы ТМС могут индуцировать эпилептические приступы.
Невозможно обеспечить посредством ТМС одновременное воздействие на разные области мозга крупномасштабной сети, восстановить активность в длиннодистантных ассоциативных трактов белого вещества (например, тракты, соединяющие фронтальные и затылочные либо фронтальные и теменные области мозга), воздействуя на них одновременно с разных концов в пределах полушария, либо в комиссуральных трактах, воздействуя одновременно на оба полушария. Кроме того, у ТМС меньшие возможности варьирования параметров магнитного поля и локализации стимуляции. Повторные сеансы ТМС могут индуцировать эпилептические приступы.
Быстрота и множественность проявления положительных терапевтических эффектов при разных по своей природе поражениях мозга дают основание полагать, что ТМС и ТМЭС изменяют функционирование сетевой структуры мозга за счет разных механизмов ПМ, воздействуя одновременно на множество разных молекул. Наиболее вероятные мишени их воздействия — белки с повышенным содержанием заряженных аминокислот. Быстрые и нарастающие от процедуры к процедуре эффекты, по-видимому, связаны с восстановлением функций нейронов, подавленных интоксикацией и отеком, предотвращением апоптоза, а также с активированием процессов П.М. При детском церебральном параличе и задержке психомоторного развития детей младшего возраста эффекты ТМЭС реализуются вне острого повреждения и связаны, возможно, с запуском процессов нейрогенеза [20]. Следует заметить, что под нейрогенезом в широком смысле следует понимать образование не только новых нейронов, но и олигодендроцитов и астроцитов, играющих активную роль в синаптической пластичности.
Современные научные технологии открывают большие возможности по регуляции ПМ и приданию ей желаемой направленности. Между отдельными индивидуумами, женщинами и мужчинами, детьми и взрослыми на макроуровне вариабельность мозга в норме проявляется не только в его объеме (массе) и в расположении извилин, но и в картинах связанности между различными его областями и плотности связей [23].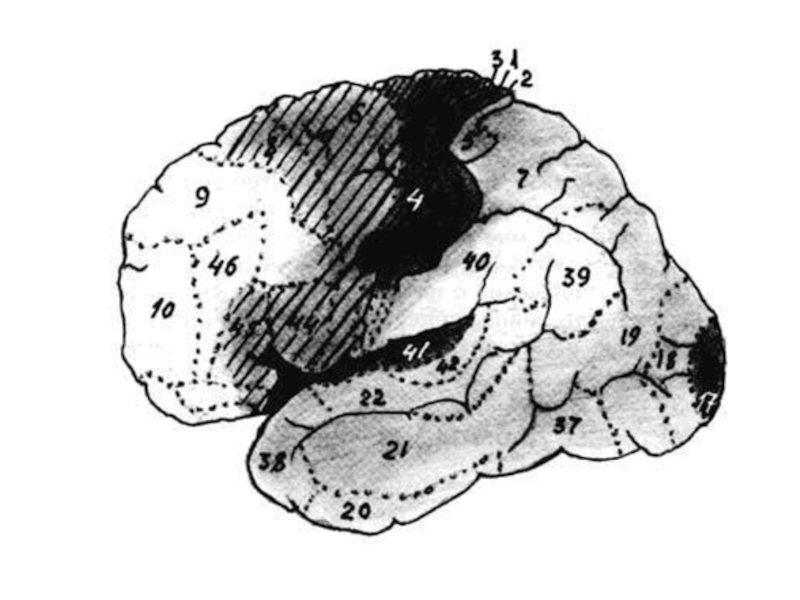 Совокупность всех связей мозга сегодня именуется как коннектом. Особую важность представляют данные о различных вариантах коннектома мозга человека. Очевидная ценность таких данных для клиники в том, что, ориентируясь на особенности коннектома пациента, можно направить ТМС или ТMЭС на быструю активацию сохраненных избыточных параллельных нервных путей и привлечь новые области мозга для восстановления утраченных функций. К примеру, известно несколько непервичных моторных кортикальных областей: премоторная кора, дополнительная моторная зона и в области, прилежащей к ней, а также ростральные, дорсальные и вентральные области поясной извилины [24]. Активация параллельных путей к спинальным мотонейронам может затрагивать как эти контралатеральные неповрежденные моторные области мозга, так и соматосенсорные области, мозжечок, базальные ганглии [25]. Анатомическая связь нейронов простирается на большее пространство, чем территория их функционального влияния, и значительная часть связей находится в состоянии тонического ингибирования, реализуемого в значительной степени, по-видимому, ГАМКергическими нейронами [26].
Совокупность всех связей мозга сегодня именуется как коннектом. Особую важность представляют данные о различных вариантах коннектома мозга человека. Очевидная ценность таких данных для клиники в том, что, ориентируясь на особенности коннектома пациента, можно направить ТМС или ТMЭС на быструю активацию сохраненных избыточных параллельных нервных путей и привлечь новые области мозга для восстановления утраченных функций. К примеру, известно несколько непервичных моторных кортикальных областей: премоторная кора, дополнительная моторная зона и в области, прилежащей к ней, а также ростральные, дорсальные и вентральные области поясной извилины [24]. Активация параллельных путей к спинальным мотонейронам может затрагивать как эти контралатеральные неповрежденные моторные области мозга, так и соматосенсорные области, мозжечок, базальные ганглии [25]. Анатомическая связь нейронов простирается на большее пространство, чем территория их функционального влияния, и значительная часть связей находится в состоянии тонического ингибирования, реализуемого в значительной степени, по-видимому, ГАМКергическими нейронами [26]. При устранении ингибирования эти молчащие связи быстро демаскируются, что лежит, по-видимому, в основе одного из быстрых механизмов ПМ и может быть названо, следуя Донохью [27], системным расширением горизонтальной связанности нейронов. Активация ипсилатеральных двигательных путей интактного полушария также может восполнить поражение контралатеральных путей. Другой быстрый механизм ПМ связан с усилением либо ослаблением существующих синаптических связей посредством долговременного потенцирования и подавления, а также с изменением порога возбудимости мембран нейронов.
При устранении ингибирования эти молчащие связи быстро демаскируются, что лежит, по-видимому, в основе одного из быстрых механизмов ПМ и может быть названо, следуя Донохью [27], системным расширением горизонтальной связанности нейронов. Активация ипсилатеральных двигательных путей интактного полушария также может восполнить поражение контралатеральных путей. Другой быстрый механизм ПМ связан с усилением либо ослаблением существующих синаптических связей посредством долговременного потенцирования и подавления, а также с изменением порога возбудимости мембран нейронов.
Перспективно использование данных по коннектому и в индукции кросс-модальной ПМ при поражении сенсорных (особенно слуховых и зрительных) представительств в мозгу, а также в стимуляции речевой системы — одной из наиболее сложных крупномасштабных систем мозга, уязвимой при инсультах и травмах и часто подверженной дизонтогенезу. Данные по коннектому и функциональной магнитной томографии позволят выявить индивидуальные особенности структурно-функциональной дезорганизации речевой системы и прицельно активировать в ней молчащие области.
Завершая обсуждение некоторых возможностей управления ПМ, нельзя обойти вниманием проблему биомаркеров при поражениях мозга. По сравнению с другими органами существующие барьеры для проникновения макромолекул из мозга в кровь резко ограничивают возможности выявления его биомаркеров. Среди немногих известных биомаркеров аполипопротеин CIII, позволяющий дифференцировать ишемический инсульт от геморрагического [28]. Применительно к поражениям при инсульте или травме мозга актуальна разработка новых вычислительных алгоритмов, позволяющих по данным нейровизуализации корректно прогнозировать варианты исхода [29].
Cтруктурно-функциональная сложность мозга проявляется на разных уровнях и в разных пространственных и временны́х шкалах. Уникальность человеческого мозга ограничивает трансляцию в клинику результатов, полученных на экспериментальных моделях, и она еще более затрудняется при включении эпигенетической регуляции. При регуляции ПМ невозможно игнорировать тесное переплетение структурно-функциональной организации мозга с ИС и сосудистой системой [5, 30], и наиболее эффективными воздействиями на ПМ и восстановление его функций, по-видимому, являются те, которые затрагивают широкий спектр молекул в разных типах клеток мозга. Такими свойствами обладают ТМС и ТMЭС, чем, вероятно, объясняется широкий спектр заболеваний, при которых они оказались эффективными, и соответственно резко возросший интерес к этим методам терапии. Использование данных по коннектому [31] и функциональной магнитно-резонансной томографии мозга может помочь точнее локализовывать эффекты ТМС и ТMЭС [22]. Стартовавшие новые крупные проекты по изучению мозга, несомненно, расширят и углубят наши представления о его сложности и откроют новые пути управления его пластичностью.
Такими свойствами обладают ТМС и ТMЭС, чем, вероятно, объясняется широкий спектр заболеваний, при которых они оказались эффективными, и соответственно резко возросший интерес к этим методам терапии. Использование данных по коннектому [31] и функциональной магнитно-резонансной томографии мозга может помочь точнее локализовывать эффекты ТМС и ТMЭС [22]. Стартовавшие новые крупные проекты по изучению мозга, несомненно, расширят и углубят наши представления о его сложности и откроют новые пути управления его пластичностью.
Конфликт интересов отсутствует.
Про нейропластичность мозга — Офтоп на vc.ru
Ответы на самые популярные вопросы из «Гугла» и «Яндекса», плюс пачка интересностей из книги Нормана Дойджа.
{«id»:181640,»url»:»https:\/\/vc. ru\/flood\/181640-quick-faq-pro-neyroplastichnost-mozga»,»title»:»Quick FAQ: \u041f\u0440\u043e \u043d\u0435\u0439\u0440\u043e\u043f\u043b\u0430\u0441\u0442\u0438\u0447\u043d\u043e\u0441\u0442\u044c \u043c\u043e\u0437\u0433\u0430″,»services»:{«facebook»:{«url»:»https:\/\/www.facebook.com\/sharer\/sharer.php?u=https:\/\/vc.ru\/flood\/181640-quick-faq-pro-neyroplastichnost-mozga»,»short_name»:»FB»,»title»:»Facebook»,»width»:600,»height»:450},»vkontakte»:{«url»:»https:\/\/vk.com\/share.php?url=https:\/\/vc.ru\/flood\/181640-quick-faq-pro-neyroplastichnost-mozga&title=Quick FAQ: \u041f\u0440\u043e \u043d\u0435\u0439\u0440\u043e\u043f\u043b\u0430\u0441\u0442\u0438\u0447\u043d\u043e\u0441\u0442\u044c \u043c\u043e\u0437\u0433\u0430″,»short_name»:»VK»,»title»:»\u0412\u041a\u043e\u043d\u0442\u0430\u043a\u0442\u0435″,»width»:600,»height»:450},»twitter»:{«url»:»https:\/\/twitter.com\/intent\/tweet?url=https:\/\/vc.ru\/flood\/181640-quick-faq-pro-neyroplastichnost-mozga&text=Quick FAQ: \u041f\u0440\u043e \u043d\u0435\u0439\u0440\u043e\u043f\u043b\u0430\u0441\u0442\u0438\u0447\u043d\u043e\u0441\u0442\u044c \u043c\u043e\u0437\u0433\u0430″,»short_name»:»TW»,»title»:»Twitter»,»width»:600,»height»:450},»telegram»:{«url»:»tg:\/\/msg_url?url=https:\/\/vc.
ru\/flood\/181640-quick-faq-pro-neyroplastichnost-mozga»,»title»:»Quick FAQ: \u041f\u0440\u043e \u043d\u0435\u0439\u0440\u043e\u043f\u043b\u0430\u0441\u0442\u0438\u0447\u043d\u043e\u0441\u0442\u044c \u043c\u043e\u0437\u0433\u0430″,»services»:{«facebook»:{«url»:»https:\/\/www.facebook.com\/sharer\/sharer.php?u=https:\/\/vc.ru\/flood\/181640-quick-faq-pro-neyroplastichnost-mozga»,»short_name»:»FB»,»title»:»Facebook»,»width»:600,»height»:450},»vkontakte»:{«url»:»https:\/\/vk.com\/share.php?url=https:\/\/vc.ru\/flood\/181640-quick-faq-pro-neyroplastichnost-mozga&title=Quick FAQ: \u041f\u0440\u043e \u043d\u0435\u0439\u0440\u043e\u043f\u043b\u0430\u0441\u0442\u0438\u0447\u043d\u043e\u0441\u0442\u044c \u043c\u043e\u0437\u0433\u0430″,»short_name»:»VK»,»title»:»\u0412\u041a\u043e\u043d\u0442\u0430\u043a\u0442\u0435″,»width»:600,»height»:450},»twitter»:{«url»:»https:\/\/twitter.com\/intent\/tweet?url=https:\/\/vc.ru\/flood\/181640-quick-faq-pro-neyroplastichnost-mozga&text=Quick FAQ: \u041f\u0440\u043e \u043d\u0435\u0439\u0440\u043e\u043f\u043b\u0430\u0441\u0442\u0438\u0447\u043d\u043e\u0441\u0442\u044c \u043c\u043e\u0437\u0433\u0430″,»short_name»:»TW»,»title»:»Twitter»,»width»:600,»height»:450},»telegram»:{«url»:»tg:\/\/msg_url?url=https:\/\/vc. ru\/flood\/181640-quick-faq-pro-neyroplastichnost-mozga&text=Quick FAQ: \u041f\u0440\u043e \u043d\u0435\u0439\u0440\u043e\u043f\u043b\u0430\u0441\u0442\u0438\u0447\u043d\u043e\u0441\u0442\u044c \u043c\u043e\u0437\u0433\u0430″,»short_name»:»TG»,»title»:»Telegram»,»width»:600,»height»:450},»odnoklassniki»:{«url»:»http:\/\/connect.ok.ru\/dk?st.cmd=WidgetSharePreview&service=odnoklassniki&st.shareUrl=https:\/\/vc.ru\/flood\/181640-quick-faq-pro-neyroplastichnost-mozga»,»short_name»:»OK»,»title»:»\u041e\u0434\u043d\u043e\u043a\u043b\u0430\u0441\u0441\u043d\u0438\u043a\u0438″,»width»:600,»height»:450},»email»:{«url»:»mailto:?subject=Quick FAQ: \u041f\u0440\u043e \u043d\u0435\u0439\u0440\u043e\u043f\u043b\u0430\u0441\u0442\u0438\u0447\u043d\u043e\u0441\u0442\u044c \u043c\u043e\u0437\u0433\u0430&body=https:\/\/vc.ru\/flood\/181640-quick-faq-pro-neyroplastichnost-mozga»,»short_name»:»Email»,»title»:»\u041e\u0442\u043f\u0440\u0430\u0432\u0438\u0442\u044c \u043d\u0430 \u043f\u043e\u0447\u0442\u0443″,»width»:600,»height»:450}},»isFavorited»:false}
ru\/flood\/181640-quick-faq-pro-neyroplastichnost-mozga&text=Quick FAQ: \u041f\u0440\u043e \u043d\u0435\u0439\u0440\u043e\u043f\u043b\u0430\u0441\u0442\u0438\u0447\u043d\u043e\u0441\u0442\u044c \u043c\u043e\u0437\u0433\u0430″,»short_name»:»TG»,»title»:»Telegram»,»width»:600,»height»:450},»odnoklassniki»:{«url»:»http:\/\/connect.ok.ru\/dk?st.cmd=WidgetSharePreview&service=odnoklassniki&st.shareUrl=https:\/\/vc.ru\/flood\/181640-quick-faq-pro-neyroplastichnost-mozga»,»short_name»:»OK»,»title»:»\u041e\u0434\u043d\u043e\u043a\u043b\u0430\u0441\u0441\u043d\u0438\u043a\u0438″,»width»:600,»height»:450},»email»:{«url»:»mailto:?subject=Quick FAQ: \u041f\u0440\u043e \u043d\u0435\u0439\u0440\u043e\u043f\u043b\u0430\u0441\u0442\u0438\u0447\u043d\u043e\u0441\u0442\u044c \u043c\u043e\u0437\u0433\u0430&body=https:\/\/vc.ru\/flood\/181640-quick-faq-pro-neyroplastichnost-mozga»,»short_name»:»Email»,»title»:»\u041e\u0442\u043f\u0440\u0430\u0432\u0438\u0442\u044c \u043d\u0430 \u043f\u043e\u0447\u0442\u0443″,»width»:600,»height»:450}},»isFavorited»:false}
Что такое нейропластичность мозга?
Нейронная пластичность — это способность мозга к восстановлению и реструктуризации. Наш биокомпьютер способен значительно меняться в зависимости от требований внешних и внутренних стимулов.
Наш биокомпьютер способен значительно меняться в зависимости от требований внешних и внутренних стимулов.
Об этом трубят уже много лет во всех СМИ, но если вы вдруг не знали, то да — нервные клетки постоянно растут, восстанавливаются, мозг без проблем выстраивает новые структуры в зависимости от своих потребностей.
Как работает нейропластичность?
Представьте, что мозг — это город. Москва, например. Она постоянно перестраивается под нужды жителей и эпохи.
Точно так же работает и наш мозг. Здания сносятся, плитка перекладывается, дороги укладываются. Единственная разница — весь бюджет 100% уходит на нужды мозга, никаких распилов на госконтрактах.
Здания сносятся, плитка перекладывается, дороги укладываются. Единственная разница — весь бюджет 100% уходит на нужды мозга, никаких распилов на госконтрактах.
Как тренировать нейропластичность?
Никак. Это не навык, это свойство мозга. Просто пользуйтесь им.
Теряется ли нейропластичность с возрастом?
Свежие исследования говорят о том, что ухудшается, но не в целом, а только в отдельных моментах. При этом есть и вторая сторона медали — некоторые способности наоборот с возрастом начинают работать лучше.
А теперь нечто совсем другое
Так как популярных вопросов про нейроплатичность не так много, то чтобы еще чуточку порадовать ваш мозг — большая подборка крутых фактов про это явление и мозг в целом. Большая часть из них — конспект основных тезисов книги «Пластичность мозга» Нормана Дойджа.
Она великолепна и крайне рекомендуется для изучения!
Сила мысли
Каждая мысль меняет мозг. Звучит с налетом эзотерики, но всё намного банальнее и довольно логично — новая информация создает новые, пусть иногда и довольно мимолетные нейронные связи.
Звучит с налетом эзотерики, но всё намного банальнее и довольно логично — новая информация создает новые, пусть иногда и довольно мимолетные нейронные связи.
Иногда это происходит очень быстро: если полностью лишить вас зрения, то уже через два дня мозг перестроится острее считывать другие сенсорные данные.
Работа мысли (и воображения) меняет ментальные карты, усиливает определённые нейронные связи. Это можно использовать для создания протезов для парализованных, а также для обучения. Например, в виртуальной реальности.
Несколько взрывающих мозг экспериментов
Две группы людей, никогда не игравшие прежде на пианино, учились исполнять последовательность нот. Затем одна группа 5 дней по 2 часа сидела перед пианино и представляла, что играет эти ноты и слушает исполнение. Другая группа действительно играла.
Затем одна группа 5 дней по 2 часа сидела перед пианино и представляла, что играет эти ноты и слушает исполнение. Другая группа действительно играла.
В результате психическая тренировка привела к тем же изменениям в двигательной системе, что и физическая. Тем, кто тренировался мысленно, потребовалось всего 2 часа дополнительной физической тренировки, чтобы достигнуть того же уровня исполнения.
Очевидно, что психическая тренировка — эффективный способ подготовки к освоению физического навыка с помощью минимальной физической тренировки
Второй эксперимент на ту же тему ещё сюрреалистичней. Люди тренировали мышцы пальца, те, кто в действительности двигал пальцем по 5 дней в неделю 4 недели подряд, увеличили мышечную массу на 30%, а те, кто делал это в воображении, увеличили массу на 22%.
Люди тренировали мышцы пальца, те, кто в действительности двигал пальцем по 5 дней в неделю 4 недели подряд, увеличили мышечную массу на 30%, а те, кто делал это в воображении, увеличили массу на 22%.
Звучит как мечта любого человека. Лежишь на диване, мечтаешь о прекрасном прессе и он качается…
Use It Or Lose It
То, чего можно достичь с помощью тренировок — это великолепно, но намного важнее помнить о принципе «не использовать — значит потерять». Если не кормить мозг чем-то новеньким (физически и психически стимулирующей активностью) время от времени, то расплатой как минимум потеря навыков и знаний, а как максимум — Альцгеймер или слабоумие.
Дабы в полной мере ощущать полноту жизни, мы должны учиться. Когда жизнь или отношения с человеком становятся слишком предсказуемыми и кажется, что больше нечему учиться, мы начинаем чувствовать беспокойство. Это своего рода протест пластичного мозга, который «скучает» без работы
Не все виды деятельности равнозначны в этом плане. Те, которые предполагают высокую концентрацию внимания: обучение игре на музыкальном инструменте, настольные игры, чтение или танцы, — снижают риск возникновения слабоумия. Например: обучение танцам, требующее заучивания новых движений, обеспечивает нам не только физическую, но и психическую стимуляцию и предполагает значительную концентрацию внимания. Менее напряженные виды деятельности, скажем, боулинг, присмотр за детьми и гольф, — не приводят к снижению риска возникновения болезни Альцгеймера.

Ближе к телу, ближе к делу
Физическому телу соответствует ментальная карта тела, где области органов расположены топографически: соседние участки тела в соседних участках мозга.
Во время обучения «карты» мозга меняются
К примеру, у пианистов карты мозга отвечающие за пальцы намного больше, чем у обычных людей. Принципы мастерства проясняются: с практикой происходит более эффективное использование нейронов, однако именно тех, которые подходят лучше всего для этой задачи, поэтому соответствующая обучению нейронная сеть («проекционная карта») сначала растёт, а затем перестаёт.
При этом критически важен фокус: долгосрочные изменения в мозге происходят только при большом внимании к задаче.
Половина мозга — не приговор
Реабилитация парализованных после инсульта больных возможна, она происходит благодаря постепенному обучению использования других участков мозга для выполнения прежних функций, проводя по пути, который они когда-то проходили в детстве.
Мозг можно сравнить со снежной горой, по которой накатывают трассы. Если вышел из строя основной путь, то можно проложить новый — это один из основных путей самоорганизации пластичного мозга.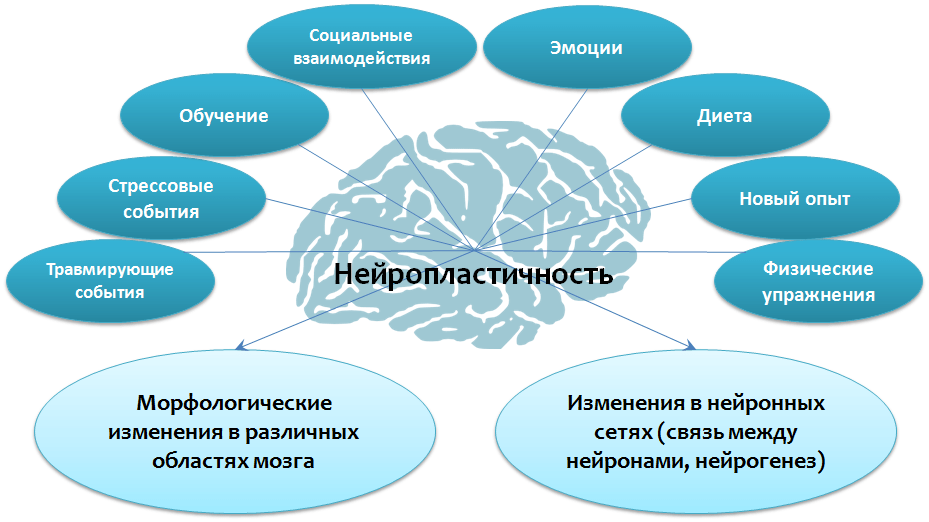
Мозг намного универсальнее, чем было принято считать вначале, как раз благодаря пластичности. Его функции не имеют жёсткой локализации — иногда другие части мозга могут взять на себя функции. Среди примеров — случаи оправившихся после инсульта или девушки, которая родилась без левого полушария, но её правое полушарие взяло на себя ряд критических функций, в том числе речь.
При инсульте движение парализуется из-за атрофии соответствующих проекционных областей в мозге, а не конкретных мышц. Интересно, что усложняется даже мысленное движение повреждённой конечностью.
Мозг — наше всё
Мы воспринимаем мир мозгом, а не органами чувств, их рецепторы — лишь ретрансляторы данных. Ощущения из одного сенсорного канала можно трансформировать в другие. Например, тактильные ощущения — в зрительные.
В эксперименте 1965 года Пол Бач-и-Рита показал, что рецепторы кожу могут заменить сетчатку глаза. Слепых от рождения людей сажали в кресло, на спинке которого было установлено 400 стимуляторов. Эти стимуляторы получали преобразованный от видеокамеры электросигнал, вибрировали в тёмных частях пространства и были неподвижны в светлых частях пространства. Через некоторое время участники эксперимента видели предметы в перспективе и даже учились узнавать предметы и людей.
Кстати, в мозге нет болевых рецепторов. Поэтому во время операции на мозге больной находится в сознании. Это активно используют для картирования мозга.
Часики тикают
У мозга есть критические периоды развития, когда мозг наиболее восприимчив к внешним стимулам и быстро растёт. Для разных навыков эти периоды разные.
Например, наилучший период для «языковых» навыков — от младенчества до 8–13 лет. Этот период — это единственное «окно», когда можно научиться мыслить сразу на двух языках.
По тому же принципу работает феномен импринтинга. Это когда, например, утята воспринимают своей мамой любой объект, который впервые видят после освобождения от скорлупы яйца.
Ученые активно ищут способы возвращать мозг в критические периоды, чтобы эффективнее учиться. Надеются делать это с помощью таблеток, но пока без особых успехов. О чём-то подобном говорил Николас Негропонте в своём TED-talk’e.
Ментальное старение происходит оттого, что портится не столько память, сколько способность к фиксированию новых событий и снижается сила и острота восприятия. Это происходит отчасти оттого, что становясь зрелыми мы перестаём учиться и пользуемся лишь тем, что уже освоили — короче, перестаём тренировать пластичность мозга. В общем, в старости нужно учить новые языки и развивать слуховую память (есть особые программы, которые позволяют «вернуться» в когнитивном развитии на 10–25 лет).
Почему тебя возбуждают чайники
Сексуальность тоже родом из мозга и чертовски пластична. Она формируется в критические периоды под влиянием внешних факторов. Тут много интересной антропологии: «В Европе романтическая любовь, со всем ее поэтическим оформлением, начала находить социальное одобрение лишь среди аристократов и при королевских дворах, да и то только в двенадцатом веке.
Как правило, это был любовный альянс между неженатым мужчиной и замужней женщиной, который обычно плохо заканчивался. Только с началом распространения идеалов, ставящих индивидуальность человека выше, идея о праве человека выбирать себе супруга действительно овладела умами людей и постепенно начала казаться совершенно естественной и неотъемлемой частью жизни».
«В Елизаветинские времена любовники были настолько одержимы запахом тел друг друга, что женщины нередко держали под мышкой очищенное яблоко до тех пор, пока оно не впитывало в себя их пот и запах. Они отдавали это «яблоко любви» своим партнерам, чтобы они могли вдыхать его аромат в их отсутствие. Мы же, напротив, применяем синтетические дезодоранты, чтобы замаскировать запах тела. Какой из этих двух подходов приобретенный, а какой природный, сказать сложно.
Например, племя масаев из Восточной Африки использует мочу коров (вызывающую у нас «естественное» отвращение) в качестве лосьона для волос, что говорит о значимости коров в их культуре. Многие склонности, которые мы считаем «природными», приобретены нами через научение и становятся для нас «второй натурой».
И почему ты возбуждаешься, когда тебя трогают за мочку уха
Из карт мозга, составленных Пенфилдом, видно, что зона гениталий расположена рядом с зоной стопы, а поскольку карта стопы больше не получает входящей информации, вполне вероятно, что ее захватывает карта гениталий, поэтому когда гениталии испытывают удовольствие, то же самое происходит в фантомной стопе. Рамачандран задумался: не может ли свойственное некоторым людям эротическое пристрастие к стопам отчасти быть вызвано близостью проекционных зон стоп и гениталий в мозге. Это позволяет объяснить и другие эротические загадки.
Один итальянский врач, доктор Сальваторе Аглиоти, сообщал: порой женщины, перенесшие мастэктомию, чувствуют сексуальное возбуждение, когда им стимулируют уши, ключицы и грудину. На карте мозга зоны этих частей тела располагаются рядом с сосками.
Наконец, известно, что некоторые мужчины с карциномой пениса, в результате которой была проведена его ампутация, чувствуют не только фантомный пенис, но и фантомные эрекции.
Незрячие читают намного быстрее зрячих
И речь не про шрифт Брайля, а про прослушивание аудиокниг. В качестве иллюстрации вот такая история:
Слепота Эммы изменила ее мозг и всю ее жизнь. Некоторые из присутствующих на ужине интересовались литературой, но Эмма, с тех пор как ослепла, прочитала гораздо больше, чем любой из нас.
Специальная машина для незрячих, разработанная компанией Kurzweil Educational Systems, монотонно читает ей вслух, делая паузы для выделения запятых, останавливаясь для выделения абзацев и повышая тон для выделения вопросов. Этот компьютерный голос звучит так быстро, что я не могу разобрать ни одного слова.
Однако Эмма постепенно училась слушать его на все большей и большей скорости, так что теперь она читает около 340 слов в минуту и занимается изучением всех классиков мировой литературы. «Я выбираю автора и читаю все, что он написал, а потом перехожу к следующему». Она прочитала всего Достоевского (это ее любимый писатель), Гоголя, Толстого, Тургенева, Диккенса, Честертона, Бальзака, Гюго, Золя, Флобера, Пруста, Стендаля и многих других.
Недавно она прочитала три романа Троллопа за один день. Однажды она спросила меня, как это может быть, что теперь она читает гораздо быстрее, чем делала это до того, как ослепла. Я предположил, что ее обширная зрительная зона коры головного мозга, которая больше не обрабатывает информацию, поступающую от органов зрения, приняла на себя обработку слуховой информации».
(Фантомная) боль
Когда человек теряет часть тела, то ментальная карта конечности вместо того, чтобы исчезнуть, начинает обрабатывать информацию с «соседнего» участка тела (в случае с рукой — с лица).
Вилейанур Субраманиан Рамачандран придумал остроумную «ампутацию фантомной конечности» посредством иллюзий, заставляя с помощью зеркального ящика почувствовать мозг, будто отсутствующая конечность двигается.
Ещё классный эксперимент Рамачандрана – если фальшивую резиновую руку расположить с рукой подопытного, оставив в видимости только фальшивую руку, а затем синхронно бить и трогать обе руки, то через несколько минут подопытному начнёт казаться, что он воспринимает ощущение удара от фальшивой руки. Она станет частью образа его тела. Можно пойти дальше и слить сенсорный образ руки с мебелью — столешницей, если повторить фокус с синхронными ударами.
Обычная боль — это тоже иллюзия мозга. Он не просто пассивно получает сигналы о боли, но контролирует их. Именно поэтому раненные на поле боя могут не чувствовать боли, а таблетки-плацебо и мамино утешение снижают её степень.
Спасибо за внимание! Надеюсь, что было интересно и полезно.
#Quick_FAQ – это серия постов, где я собираю все самые часто задаваемые вопросы по определенной теме, а потом подбираю для них релевантные, научно-доказанные и рабочие ответы.
Больше интересного и полезного про биохимию мозга, обучение и то, как настраивать себя на продуктивные будни можно найти на моём телеграм-канале. Подписывайтесь! Буду рад увидеть вас среди своих читателей.
Функциональная межполушарная асимметрия и пластичность мозга
Иллариошкин С.Н., Фокин В.Ф.
М.: ФГБУ «НЦН» РАМН, 2012.
Два года, прошедшие со времени проведения предыдущей конференции, посвященной актуальным вопросам функциональной межполушарной асимметрии и нейропластичности, показали, что мировая тенденеция развития нейронаук в значительной мере опирается на фундамент этих двух научных дисциплин. Современные наукометрические методы позволили выявить, что эти два научных направления являются ярко выраженными мировыми трендами, как по количеству публикаций, так и по числу цитированных работ. Исторически в России сложились сильные научные школы по изучению функциональной межполушарной асимметрии и нейропластичности. На этой конференции большинство из них представлены в серии докладов и отражены в материалах конференции. Проблема функциональной межполушарной асимметрии по своей сути является мультидисциплинарной, она объединяет специалистов, работающих в различных областях биологии, медицины, психологии и, конечно, нейропластичности. Хотел бы отметить консолидирующую роль журнала «Асимметрия» в развитии этого научного направления. Проблема нейропластичности также является мультидисциплинарной и охватывает, как и предыдущая, широкий круг тем — от сугубо экспериментальных исследований до клинической медицины. Проблемы функциональной межполушарной асимметрии и нейропластичности в ряде областей пересекаются. Это, прежде всего, исследование онтогенеза, а также возрастного развития и старения мозга. Кроме того, общим, является и целый ряд клинических направлений, особенно тех, в которых патологический процесс затрагивает преимущественно одно полушарие.
Сейчас остро ставится вопрос об интеграции отечественной науки с общемировой. Поэтомусудовлетворениемхочетсяотметить.чтозначительная часть исследований, представленных в материалах конференции, выполнена на высоком профессиональном уровне, во многом сопоставимом с уровнем работ наших зарубежных коллег. Кроме того приятным для нас является факт участия в работе конференции ряда ведущих иностранных ученых из ближнего и дальнего зарубежья.
Есть все основания полагать, что настоящая конференция, так же как и предыдущие, станет серьезным шагом вперед в развитии исследований по функциональной межполушарной асимметрии и нейропластичности.
Заместитель директора ФГБУ «НЦН» РАМН,
Руководитель Отдела исследований мозга
Иллариошкин С.Н.
Пластичность мозга и современные реабилитационные технологии Текст научной статьи по специальности «Клиническая медицина»
Пластичность мозга и современные реабилитационные технологии
Л.А. Черникова
НИИ неврологии РАМН, г. Москва
В работе обсуждаются основные достижения в области изучения механизмов нейропластичности с помощью современных методов нейровизуализации и, прежде всего, функциональной МРТ. Подчеркивается роль в этих процессах различных афферентных воздействий. Современные нейрореабилитационные технологии, такие как С1-терапия, система «ЮСОИЛТ», робототерапия и др., рассматриваются как источники усиленной целенаправленной афферентации. Приводятся данные о возможности применения нервно-мышечной электростимуляции в первые часы после инсульта. Обсуждается уникальная методика внутриглоточной электростимуляции при лечении дисфагий различной этиологии. Приводятся данные об эффективности транскраниальной электростимуляции при лечении центрального постинсультного болевого синдрома. Представлены возможности метода биоуправления, организованного по электромиограмме, при обучении больных точностному схвату — одному из основных двигательных навыков руки. Детально обсуждаются особенности обучения различным постуральным задачам методом биоуправления по статокинезиграмме у больных с постинсультными гемипарезами, болезнью Паркинсона и спиноцеребеллярными атаксиями. Представлены данные об эффективности альфа-стимулирующего тренинга у больных с центральным постинсультным болевым синдромом с преобладанием в клинической картине аффективных расстройств. В заключение обсуждаются перспективы одной из самых интересных современных реабилитационных технологий — технологии, основанной на виртуальной реальности.
Ключевые слова: нейропластичность, афферентные воздействия, новые реабилитационные технологии.
Одним из наиболее выдающихся достижений в области фундаментальных наук второй половины ХХ века являются результаты исследований пластичности головного мозга, доказавшие, что кора взрослого человека способна к значительным функциональным перестройкам и что решающая роль в этой реорганизации принадлежит афферентной информации. Эти достижения позволили по-новому взглянуть на роль нейрореабилитации, которая в настоящее время окончательно теряет свой статус Золушки в мире неврологии и начинает занимать одно из приоритетных направлений в клинической медицине [15].
Предположение о возможности ремодулирования невральных кортикальных соединений под влиянием тренировки одним из первых высказал Б. ИеЪЪ [26] еще в 40-е годы прошлого века. Но уже начиная с середины 60-х годов стали появляться многочисленные экспериментальные исследования, демонстрирующие химические и анатомические механизмы пластичности головного мозга у взрослых животных [14, 51, 29]. Наиболее полно возможность реорганизации кортикальных полей под влиянием сенсорного входа была продемонстрирована только в середине 80-х годов М. МегсешсИ с соавторами [32, 38, 39] в опытах на обезьянах, в которых удалось показать увеличение области сенсомоторной коры под влиянием контролируемой тактильной стимуляции и уменьшение коркового представительства кисти и соответствующее расширение представительства соседних сегментов руки — предплечья и плеча при ампутации кисти. Тогда же эти авторы впервые высказали предположение об актуальности полученных данных для реабилитации больных после инсульта.
Действительно, сохранность двигательного представительства той или иной части тела обеспечивается афферентным потоком от нее. Так, при постоянной афферентации с кисти, например у пианистов или слепых, читающих при помощи шрифта Брайля, увеличивается представительство заинтересованных зон [46, 22]. Более того, в исследовании А. РаБсиа1-Ьеопе с соавторами [47] при использовании метода транскраниальной магнитной стимуляции на здоровых добровольцах было показано, что временная функциональная реорганизация областей кортикального представительства может стать обычным явлением в каждодневной жизни при обучении.
В настоящее время благодаря методам функциональной визуализации появилась возможность изучать процессы нейропластичности, развивающиеся в головном мозге при его повреждении. Одно из первых исследований в этой области было выполнено Я. №ёо и в. МПИкеп [45], которые, произведя частичное повреждение моторной коры обезьяны, показали, что без последующей тренировки представительство пораженной конечности уменьшалось в размере. Но если паретичную конечность тренировать, то ее двигательное представительство увеличивалось, распространяясь на области, формально отвечающие за представительство плеча и локтя. Таким образом, тренинг оказывает прямое влияние на реорганизацию коры мозга с вовлечением прилежащей к инфаркту непораженной моторной коры. Во многих исследованиях показано, что в процессе обучения новым двигательным навыкам участвуют разные мозговые структуры в зависимости от характера тренируемого движения, такие как префронтальная, париетальная и соматосенсорная кора, премоторная, первичная и дополнительная моторная кора, таламус, мозже-
чок как на стороне поражения, так и на здоровой стороне [21, 24, 30, 52]. Так, в исследовании R. Seitz [53] с соавторами выявлено последовательное включение мозжечка и премоторной коры при обучении воспроизведения определенной траектории движения. Интересные результаты получены в работе H. van Mier с соавторами [40]: оказалось, что при первичном обучении двигательным навыкам в руке и при последующей двигательной тренировке активируются разные комплексы мозговых структур.
В настоящее время значительная роль в восстановлении нарушенных функций при поражении первичных моторных зон и пирамидных трактов в бассейне средней мозговой артерии отводится более дистально расположенным от первичной моторной коры зонам, а именно премоторной коре [41, 42], которая обладает рядом свойств, позволяющих ей взять на себя функцию первичной моторной коры при ее повреждении. Это прежде всего высокая видоизменяемость ее нейронов при обучении, прямые проекции пирамидных клеток V слоя на спинальные интернейроны и альфа-мотонейроны, наконец, соматотопическая организация, аналогичная такой же в первичной моторной коре.
Каковы же механизмы нейропластичности?
Согласно современным представлениям, как при модификации синаптических соединений, так и при реорганизации кортикальных полей у взрослых вовлекается прежде всего феномен долговременной потенциации (ДП), заключающийся в длительном увеличении нейронной активности (синтезе новых белков и развитии новых синаптических соединений) определенной структуры после тетани-зации ее афферентного входа. Этот феномен лежит в основе процессов обучения и памяти. Для выработки ДП необходимы три условия:
1. содружественность активирования нескольких нервных волокон;
2. ассоциативность активирования пресинаптических волокон и постсинаптических клеток;
3. специфичность механизма ДП только к активированному пути.
Механизм ДП лежит в основе таких синаптических изменений, как:
1. восстановление синаптической передачи по мере разрешения локального отека, сдавливающего тело нейрона или аксон, в результате чего возникает невральный шок;
2. денервационная гиперчувствительность, которая имеет место при разрушении окончания пресинаптического аксона. На клетке-мишени развиваются новые рецепторы в ответ на медиатор, высвобождающийся из других близлежащих аксонов. Примером может служить денер-вационная гиперчувствительность в постсинаптической мембране клеток стриатума, которая развивается при гибели нейронов черной субстанции;
3. синаптическая гиперэффективность, которая развивается при разрушении только некоторых ветвей пресинап-тического аксона. Оставшиеся терминали получают весь медиатор, который в норме распределяется среди всех
окончаний. В результате на оставшиеся окончания аксона поступает большее количество медиатора, чем в нормальных условиях. Механизм ДП, по-видимому, превращает молчащие синапсы в активные путем структурных изменений этих синапсов при самом активном участии АМРА- и NMDA-глютаматных рецепторов, а также Са2+ и Na+ каналов. Надо сказать, что глютамат играет решающую роль в механизмах нейропластичности. В ряде экспериментальных работ показано, что блокирование NMDA-рецепторов глютамата предотвращает реорганизацию кортикальных карт первичной соматосенсорной коры.
Что же касается аксональных изменений, то известно, что регенеративный спраутинг (возобновление роста пораженного аксона) типичен только для периферической нервной системы, поскольку повреждения аксонов в ЦНС носят необратимый характер; это обусловлено отсутствием фактора роста нервов (NGF), вырабатываемого шваннов-скими клетками, торможением роста за счет олигодендро-цитов и фагоцитарной активностью микроглии. Вместе с тем существование другого вида спраутинга (коллатерального) в структурах головного мозга (в рубро-спинальных нейронах красного ядра) впервые было показано в уникальных экспериментальных исследованиях N. Tsukahara еще в середине 1980-х годов [23]. Затем в ряде работ H. Asanuma с соавторами были обнаружены явления спраутинга и формирование новых синапсов в моторной коре за счет входа из соматосенсорной коры после уменьшения таламического входа [11, 33, 43]. Наконец, в одной из последних работ R. Nudo с соавторами [45], выполненных на обезьянах, было показано изменение траектории внутрикортикальных аксонов вентральной премоторной зоны вблизи очага ишемии, расположенного в первичной моторной зоне у обезьян, проходивших целенаправленный тренинг руки, и отсутствие этого явления у обезьян контрольной группы, которые не тренировались. Эти данные еще раз доказывают наличие феномена аксонального коллатерального спраутинга в мозговой коре.
Таким образом, в многочисленных экспериментальных и клинических исследованиях было выявлено, что в активизации механизмов нейропластичности ЦНС важную роль играют различные методы усиления афферентного входа, которые составляют основу восстановительной терапии больных с двигательными нарушениями. Эти данные, несомненно, послужили значительным толчком к развитию новых технологий в области двигательной реабилитации.
Кинезитерапия________________________________________
Наряду с традиционно применяемыми методами ЛФК, состоящими из классических лечебно-гимнастических приемов и направленными на выработку отдельных движений, в современной кинезитерапии все шире внедряются новые подходы, ориентированные на тренировку и обучение определенной двигательной задачи (task-oriented approach) [16, 56]. Такие подходы основаны на интенсификации лечебных методик, внедрении специальных робототехнических устройств.
К настоящему времени накоплен достаточно большой материал об эффективности метода интенсивной тренировки паретичной руки у больных с легкими или умерен-
Том 1. №2 2GG7
рис. 1: Форсированная методика кинезитерапии — Сопз1га1пЫпЛсе(1 (С1)-терапия
Обучение двигательным навыкам в паретичной руке стимулируется путем фиксации здоровой руки в течение 2 недель по 5 часов в день.
&
рис. 2: Система «LOCOMAT»
ными парезами, при котором здоровая рука остается жестко фиксированной в течение 5-6 часов в день, в то время как паретичная рука усиленно тренируется (constraint-induced movement therapy — CI therapy) [57] (рис. 1). В настоящее время эффективность этой технологии в реабилитации больных с легкими и умеренными парезами при разной давности инсульта, начиная с 3 мес. и более, не вызывает сомнения [34, 48, 49]. Улучшение двигательных функций руки, наблюдаемое при использовании CI therapy, нашло подтверждение в последнее время в исследованиях с применением транскраниальной магнитной стимуляции, функциональной МРТ (фМРТ) и однофотонной эмиссионной компьютерной томографии (ОФЭКТ), выявивших
процессы реорганизации коркового представительства руки не только в здоровом, но и в пораженном полушарии под влиянием этой терапии [18, 35, 50].
В качестве еще одного перспективного способа интенсификации кинезитерапии можно рассматривать методику тренировки ходьбы с использованием бегущих дорожек с поддерживающими вес тела системами, которая признана в качестве самой эффективной технологии восстановления навыка ходьбы для больных с постинсультными гемипарезами [12, 27]. В последние годы рассматриваемые системы были дополнены компьютеризированными роботами-ор-тезами, которые вначале обеспечивают пассивные движения в нижних конечностях, имитируя шаг (система «ЬОСОМАТ» фирмы Иосоша) [19], а по мере восстановления движений доля активного участия больного в локомо-ции увеличивается (рис. 2). По оценке специалистов, такая система прежде всего облегчает работу инструкторов кине-зитерапии и особенно эффективна у больных с нижней параплегией. В настоящее время получены обнадеживающие результаты применения этих систем и у больных с постинсультными гемипарезами [36].
Большой интерес представляет использование робототехнических устройств для восстановления функций руки, особенно у больных с грубыми парезами [13, 28, 59]. Одна из целей робототерапии заключается в преодолении патологических мышечных синергий, возникающих при попытке больного совершить какое-либо произвольное движение. Например, робот-ортез, фиксируемый на паре-тичной руке больного, запрограммирован таким образом, что он препятствует появлению сгибательной синергии в руке во время произвольных движений. Тренировка с помощью этого робота-ортеза в течение 8 недель (по 3 раза в неделю) приводит к значительному уменьшению выраженности синергии и увеличивает функциональные возможности руки.
Электростимулирующая терапия________________________
Другим методом реабилитации, который широко используется при центральном парезе, является метод нервномышечной электростимуляции (НЭС). Известно, что тренировочный эффект НЭС связан как с непосредственной активизацией больших мотонейронов а-типа, так и с облегчающими эффектами со стороны кожных афферен-тов на эти мотонейроны; он сравним лишь с тренирующим эффектом произвольных сокращений очень высокой интенсивности. Однако в отличие от активных физических упражнений, оказывающих прямые активирующие влияния на сердечно-сосудистую и дыхательную системы, при НЭС эти влияния минимальны и носят преимущественно локальный характер [9]. Указанное обстоятельство послужило основанием для начала в Институте неврологии РАМН цикла исследований по изучению возможности использования НЭС в острейший период инсульта (в первые часы после развития заболевания) [6]. Проведенные работы показали, что применение НЭС паретичной руки уже в первые часы после развития инсульта (стимулируются разгибатели кисти и пальцев паретичной руки по 20 мин. дважды в день в течение 3 недель) весьма положительно влияет на двигательное и функциональное восстановление больных, не ухудшая при этом состояние мозгового кровотока (по данным перфузионно-взвешенной МРТ) и не увеличивая зону инфаркта (по данным диффузионновзвешенной МРТ). Кроме того, при использовании фМРТ
рис. 3: Проведение процедуры внутриглоточной электростимуляции
во время НЭС паретичной руки было показано, что НЭС вызывает целенаправленный афферентный поток, приводящий к появлению очагов активации коры мозга на стороне поражения в периинфарктной зоне [58].
В Институте неврологии РАМН был разработан и внедрен в клинику уникальный метод внутриглоточной электростимуляции (рис. 3) у больных с нарушениями глотания различного генеза. Использование эндоскопического исследования до начала лечения позволило уточнить показания и противопоказания для применения данного метода лечения дисфагии. В настоящее время продолжаются исследования по разработке комплексной терапии с включением внутриглоточной электростимуляции и специальной методики лечебной гимнастики у больных с постин-сультной дисфагией [17, 54]. Предварительные результаты свидетельствуют о высокой эффективности такой терапии с точки зрения восстановления (хотя бы частичного) функции глотания и связанного с этим заметного улучшения качества жизни больных.
В последние годы все большее внимание уделяется методу транскраниальной электростимуляции (ТЭС), под которой подразумевается неинвазивное электрическое воздействие (прямоугольные импульсы частотой 60-80 Гц) на мозг через кожные покровы головы, избирательно активирующее эндорфинергические и серотонинергические структуры защитных систем мозга. Основные эффекты ТЭС-терапии: обезболивание, ускорение процессов эаживления, нормализация кровяного давления, лечение абстинентных синдромов, стимуляция иммунитета, нормализация психофизиологического статуса и некоторые другие [3]. В Институте неврологии ТЭС с успехом применяется для лечения центрального постинсультного болевого синдрома. Кроме того, было показано, что у больных с данным синдромом ТЭС значительно снижает выраженность гиперпатии и степень гиперкинезов [5].
Функциональное биоуправление с обратной связью
Метод функционального биоуправления с помощью обратной связи по различным физиологическим параметрам занимает особое место среди новейших реабилита-
рис. 4: Тренировка точностного схвата методом биоуправления с обратной связью по электромиограмме
ционных технологий. В основе этого метода лежит активное обращение к личности пациента и использование обратной связи как источника дополнительной информации для пациента о результативности выполнения отдельных действий, целенаправленного движения или поведения в целом.
Основная цель метода биоуправления с обратной связью заключается в повышении уровня осознания и произвольного управления как обычно не осознаваемых (непроизвольных), так и произвольно контролируемых физиологических процессов — сначала путем контролирования внешних сигналов, а затем путем сознательного регулирования внутреннего физиологического состояния или усвоения такого типа поведения, которое будет предотвращать возникновение симптомов, устранять или ослаблять их вскоре после возникновения. Можно сказать, что биоуправление, сопровождающееся широким спектром сигналов, пояснениями и обучением пациента, восполняет недостаток информации в процессе лечения.
В Институте неврологии РАМН метод биоуправления, организованный по электромиограмме, используется при тренировке точностного схвата у больных с постинсульт-ными гемипарезами [1, 10, 55] с помощью аппаратно-программного комплекса «БОС-ЛАБ», разработанного в Институте молекулярной биологии и биофизики СО РАМН (Новосибирск). В качестве сигнала обратной связи используется электрическая активность мышц возвышения большого пальца паретичной руки (рис. 4). Обучение осуществляется по специально разработанному протоколу, согласно которому периоды тренировки с использованием зрительной обратной связи по ЭМГ чередуются с воспроизведением аналогичного по силе мышечного напряжения «по памяти», без предъявления сигнала обратной связи. После курса тренировки у больных наблюдается тенденция к уменьшению амплитуды отклонения от заданного уровня тренировки, что можно расценивать как повышение точности воспроизведения дозированного мышечного напряжения. Кроме того, наблюдалось снижение амплитуды мышечной активности в фазу расслабления и достоверное увеличение максимально возможного сокращения группы мышц thenar в фазу произвольного максимального мышечного сокращения у здоровых и больных лиц.
рис.5: Функциональное биоуправление по статокинезиграмме (баланс-тренинг)
рис. 6: Альфа-стимулирующий тренинг
Одним из интересных направлений применения технологий, основанных на использовании обратной связи по различным физиологическим параметрам, является активно разрабатываемый в Институте неврологии РАМН метод биоуправления, в котором в качестве сигнала обратной связи используются параметры проекции центра давлений (ЦД) на плоскость опоры (рис. 5). Данный метод позволяет обучать больного произвольно контролировать перемещение ЦД без потери равновесия в ходе специальных компьютерных стабилографических игр. Метод осуществляется на компьютерно-стабилографическом комплексе «Ста-билан», разработанным ОКБ «РИТМ» (г. Таганрог) со специальным пакетом прикладных программ. По условию игры больной должен посредством произвольных перемещений корпуса относительно стоп совмещать ЦД, представленный на экране в виде курсора, с мишенью и далее перемещать ее в определенном направлении в зависимости от игрового задания.
Проведенные исследования показали, что тренировка методом биоуправления, организованного по статокинези-грамме, отражающей перемещение ЦД на плоскости опоры, способствует уменьшению асимметрии и повышению устойчивости вертикальной позы — как статической, так и динамической, при этом наблюдается уменьшение степени пареза в ноге, восстановление глубокой чувствительности, повышение функциональных возможностей больного в целом [8]. Кроме того, было выявлено, что включение данного метода в комплексную реабилитацию больных с постинсультными гемипарезами способствует достоверному улучшению параметров ходьбы, и прежде всего уменьшению асимметрии шага [4]. Дальнейшие исследования показали, что наличие корковых механизмов контроля позы и обучения произвольному управлению ЦД не подлежит сомнению. В последнем принимают участие различные корковые зоны. Моторная, премоторная и париетальная области участвуют как в когнитивной, так и в координационной фазе формирования нового позного навыка. Максимальный дефицит обучения возникает при комбинированных поражениях моторной и теменной или моторной, премоторной и теменной областей. Дефицит сенсорной интеграции и схемы тела в пространстве при поражении теменной области является одним из основных ограничивающих факторов и не может быть преодолен в процессе обучения [7]. Кроме того, было показано, что обучение общей стратегии управления ЦД в большей степени нарушено у больных болезнью Паркинсона и спино-церебеллярными атаксиями, чем у больных с постинсульт-ными гемипарезами. В то же время обучение тонкой поз-ной координации наиболее успешно у больных с болезнью Паркинсона по сравнению с больными с постинсультными гемипарезами и спиноцеребеллярными атаксиями. Предполагается, что механизмы участия нигро-стриарной, кор-тико-спинальной систем и мозжечка в обучении произвольному контролю позы частично являются общими, а частично различаются [31].
В настоящее время в институте начаты исследования по изучению возможности применения биоуправления, организованного по альфа-ритму, в комплексной терапии больных с ЦПИБС (рис. 6). Известно, что ЦПИБС сопровождается нарушением высших психических функций, развитием выраженных аффективных расстройств и изменениями личности астенического и депрессивно-ипохондрического типа, что замедляет функциональное восстановление пациента. В основу работ по использованию в качестве регулируемого параметра биоуправления показателей биоэлектрической активности мозга легли наблюдения за изменениями альфа-ритма у здоровых людей в зависимости от их функционального состояния. Было показано, что между различными психическими показателями (такими, как память, внимание, тревожность) и данными электроэнцефалограммы (ЭЭГ) существует определенная зависимость [25, 44]. Эти работы послужили толчком к широкому клиническому применению биоуправления по параметрам ЭЭГ (нейробиоуправление) при различных органических и функциональных заболеваниях ЦНС -эпилепсии, нарушениях сна, синдроме нарушения внимания, депрессии, травмах головного мозга, инсульте, синдроме хронической усталости, различных болевых синдромах. Проведенные исследования продемонстрировали, что альфа-стимулирующий тренинг показан больным с ЦПИБС, для которых характерны интактность перцептивно-гностической деятельности, доминирование собствен-
но депрессивного аффекта в структуре аффективных расстройств и при этом имеет место сохранность способности к обучению, наличие терапевтической мотивации. Отметим, что альфа-стимулирующий тренинг оказывает более значительное влияние на аффективную составляющую боли [2, 5].
Виртуальная реальность_______________________________
Одной из наиболее интересных и перспективных в современной нейрореабилитации является технология, основанная на виртуальной реальности (ВР), технической основой которой служат компьютерное моделирование и компьютерная имитация, а также ускоренная трехмерная визуализация, позволяющие реалистично отображать на экране движение. Несомненные достоинства этой технологии — возможность достижения большей интенсивности тренировок на фоне усиления обратной сенсорной связи, создание индивидуального виртуального пространства для каждого больного в соответствии с его двигательными особенностями, а также взаимодействие с виртуальными объектами внутри этих пространств. Для реализации ВР необходимы следующие аппаратные средства: монитор, мышь с пространственно управляемым курсором или джойстиком, иногда применяются виртуальные шлемы с дисплеями или со стереоскопическими очками, «цифровые перчатки», которые обеспечивают тактильную обратную связь с пользователем. В настоящее время технологии ВР широко применяются для тренировки локомоции и постуральной устойчивости [37]. С этой целью создаются виртуальные комнаты, коридоры, залы. Пример одного из
А
виртуальных коридоров представлен на рис. 7 (наблюдение К.И. Устиновой). В настоящее время получены убедительные свидетельства кортикальной реорганизации соответствующих зон мозга (по данным фМРТ) у больных с постинсультными гемипарезами давностью более 1 года при использовании ВР-технологий, направленных на тренировку ходьбы [60].
В заключение необходимо подчеркнуть, что благодаря появлению новых функциональных методов нейровизуализации, прежде всего фМРТ, открылись уникальные возможности как для изучения нейропластических процессов, происходящих в различных структурах мозга под влиянием тех или иных реабилитационных технологий, так и для усовершенствования этих методов восстановительной терапии и создания новых технологий. Огромное значение имеют исследования, в которых продемонстрированы кортикальная реорганизация в отдаленные периоды после инсульта — спустя 1 год и больше — на фоне применения целенаправленных интенсивных тренировок, приводящих к выработке новых двигательных навыков. Вместе с тем не следует забывать, что нейропластичность как постоянный физиологический фактор, действующий в постинсультном периоде, участвует также в формировании и реализации ряда патологических симптомов — спастичности, повышенной рефлекторной активности и хронического болевого синдрома, появление которых также может быть спровоцировано усиленным афферентным потоком. В связи с этим важнейшей задачей современной нейрореабилитации как науки является изучение возможности адекватного управления нейропластическими процессами с помощью различных средств восстановительной медицины.
Б
рис. 7: Технология виртуальной реальности: А — виртуальный коридор, Б — пациент в стереоскопических очках
Список литературы
1. Гусарова М.В., Черникова Л.А., Ланская Л.Д., Иоффе М.Е. Применение метода биоуправления с обратной связью по электро-миограмме при тренировке точностного схвата у больных с постинсультными гемипарезами. В кн.: Медицинская реабилитация пациентов с заболеваниями и повреждениями опорно-двигательной и нервной систем. М., 2004: 367-369.
2. Ланская Л.Д., Сашина М.Б., Гусарова М.В., Черникова Л.А. Эффекты альфа-стимулирующего биоуправления у больных с центральной постинсультной болью. В кн.: Медицинская реабилитация пациентов с заболеваниями и повреждениями опорнодвигательной и нервной систем. М., 2004: 383-385.
3. Лебедев В.П. Транскраниальная электростимуляция: новый подход (экспериментально-клиническое обоснование и аппаратура). В кн.: Лебедев В.П. (ред.). Транскраниальная электростимуляция. Экспериментально-клинические исследования. Сборник статей. С.Пб., 2001: 22-38.
4. Майорникова С.А., Козырева О.В., Черникова Л.А. Особенности комплексной методики восстановления функции ходьбы у больных с постинсультными гемипарезами. Леч. физкульт. и массаж 2006; 8: 3-6.
5. Сашина М.Б., Черникова Л.А., Кадыков А.С. Постинсультные болевые синдромы. Атмосфера. Нервные болезни 2004; 3: 25-27.
6. Умарова Р.М., Черникова Л.А., Танашян М.М., Кротенкова М.В. Нервно-мышечная электростимуляция в острейший период ишемического инсульта. Вопр. курортологии, физиотер. и леч. физкультуры 2005; 4: 6-8.
7. Устинова К.И., Черникова Л.А., Иоффе М.Е., Слива С.С. Нарушения обучения произвольному контролю позы при корковых поражениях различной локализации: к вопросу о корковых механизмах регуляции позы. Журн. высшей нервной деятельности 2000; 3: 421-433.
8. Черникова Л.А., Устинова К.И., Иоффе М.Е. и др. Биоуправление по стабилограмме в клинике нервных болезней. Бюллетень СО РАМН 2004; июль-сентябрь: 85-91.
9. Черникова Л.А., Шарыпова Т.Н., Разинкина Т.П., Торопова Н.Г. Влияние нервно-мышечной электростимуляции на мышечный кровоток у больных с постинсультными гемипарезами. Физиотерапия, бальнеология и реабилитация 2003; 3: 23-26.
10. Шестакова М. В., Ланская Л. Д., Билименко А. Е. и др. Обучение произвольному контролю ЭМГ со зрительной обратной связью в норме и у больных с постинсультными гемипарезами: роль зрительной и проприоцептивной афферентации. В сб.: Мат-лы международного симпозиума «Механизмы адаптивного поведения», посвященного 80-летию организации Института физиологии им. И.П. Павлова РАН. М., 2005: 6.
11. Asanuma H, Mackel R. Direct and indirect sensory input pathways to the motor cortex; its structure and function in relation to learning of motor skills. Jpn. J. Physiol. 1989; 39: 1-19.
12. Barbeau H., Visintin M. Optimal outcomes obtained with body-weight support combined with treadmill training in stroke subjects. Arch. Phys. Med. Rehabil. 2003; 84: 1458-1465.
13. BeerR.F., Dewald J.P., Dawson M.L., Rymer W.Z. Target-dependent differences between free and constrained arm movements in chronic hemiparesis. Exp. Brain Res. 2004; 156: 458-470.
14. Bennett E.L., Diamond M.C., Krech D., Rosenzweig M.R. Chemical and anatomical plasticity of brain. Science 1964; 146: 610-619.
15. Bracewell R.M. Stroke: neuroplasticity and recent approaches to rehabilitation. J. Neurol. Neurosurg. Psychiatry 2003; 74: 1465-1470.
16. Carr J.H., Shepherd R.B. Motor relearning programme for stroke. Rockville: Aspen Publications, 1983.
17. Chernikova L., Avdjunina I., Savizkaya N. et al. Effectiveness of interpharyngeal electrostimulation in patients with poststroke dysphagia. Neurologie & Rehabilitation. 2004; 4: 46.
18. Chouinard P.A., Leonard G., Paus T.Changes in effective connecti-
vity of the primary motor cortex in stroke patients after rehabilitative therapy. Exp. Neurol. 2006; 201: 375-387.
19. Colombo G., Hostettler P. Der Lokomat-eine angetriebene Geh-Orthese. Med. Orth. Tech. 2000; 120: 178-181.
20. Dancause N., Barbay S., Frost S.B. et al. Extensive cortical rewiring after brain injury. J. Neurosci. 2005; 25: 10167-10179.
21. Deiber M.P., Ibanez V., Honda M. et al. Cerebral processes related to visuomotor imagery and generation of simple finger movements studied with positron emission tomography. Neuroimage 1998; 7: 73-85.
22. Elbert T, Pantev C., Wienbruch C. et al. Increased cortical representation of the fingers of the left hand in string players. Science 1995; 270: 305-307.
23. Fujito Y., Watanabe S., Kobayashi H, Tsukahara N. Promotion of sprouting and synaptogenesis of cerebrofugal fibers by ganglioside application in the red nucleus. Neurosci. Res. 1985; 2: 407-411.
24. Goerres G.W., Samuel M., Jenkins, I.H., Brooks, D.J. Cerebral control of unimanual and bimanual movements: a PET study. Neuroreport 1998; 9: 3631-3638.
25. Hardt J.V., Kamiya J. Anxiety change through electroencephalo-graphic alpha feedback seen only in high anxiety subjects. Science 1978; 201: 79-81.
26. Hebb D.O. The effects of early experience on problem solving at maturity. Am. Psychol. 1947; 2: 737-745
27. Hesse S., Konrad M., Uhlenbrock D. Treadmill walking with partial body weight support versus floor walking in hemiparetic subjects. Arch. Phys. Med. Rehabil. 1999; 80: 421-427.
28. Hogan N., Krebs H.I., Rohrer B. et al. Motions or muscles? Some behavioral factors underlying robotic assistance of motor recovery. J. Rehabil. Res. Dev. 2006; 43: 605-618.
29. Holloway R.L. Dendritic branching: some preliminary results of training and complexity in rat visual cortex. Brain Res. 1966; 2: 393-396.
30. Honda M., Deiber M.P., Ibanez V. et al. Dynamic cortical involvement in implicit and explicit motor sequence learning. A PET study. Brain 1998; 121: 2159-2173.
31. Ioffe M.E., Ustinova K.I., Chernikova L.A., Kulikov M.A. Supervised learning of postural tasks in patients with poststroke hemiparesis, Parkinson’s disease or cerebellar ataxia. Exp. Brain Res. 2006; 168: 384-394.
32. Jenkins W.M., Merzenich M.M. Reorganization of neocortical representations after brain injury: a neurophysiological model of the bases of recovery from stroke. Progr. Brain Res. 1987; 71: 249-266.
33. Keller A., Arissian K., Asanuma H. Formation of new synapses in the cat motor cortex following lesions of the deep cerebellar nuclei. Exp. Brain Res. 1990; 80: 23-33.
34. Kopp B., Kunkel A., Muhlnickel W. et al. Plasticity in the motor system related to therapy-induced improvement of movement after stroke. Neuroreport 1999; 10: 807-810.
35. Liepert J. Motor cortex excitability in stroke before and after constraint-induced movement therapy. Cogn. Behav. Neurol. 2006; 19: 41-47.
36. Mauritz K.H. Gait training in hemiparetic stroke patients. Eura Medicophys. 2004; 40: 165-178.
37. Merians A.S., Jack D., Boian R. et al. Virtual reality-augmented rehabilitation for patients following stroke. Phys. Ther. 2002; 82: 898-915.
38. Merzenich M.M., Kaas J.H., Wall J.T. et al. Topographic reorganization of somatosensory cortical areas 3b and 1 in adult monkeys following restricted deafferentation. Neuroscience 1983; 8: 33-55.
39. Merzenich M.M., Nelson R.J., Stryker M.P. et al. Somatosensory cortical map changes following digit amputation in adult monkeys. J. Comp. Neurol. 1984; 224: 591-605.
40. Mier H., Tempel L.W., Perlmutter J.S. et al. Changes in brain activity during motor learning measured with PET effects of hand of performance and practice. J. Neurophysiol. 1998; 80: 2177-2199.
41. Miyai I., Suzuki T., Kang J. et al. Middle cerebral artery stroke that
includes the premotor cortex reduces mobility outcome. Stroke 1999; 30: 1380-1383.
42. Miyai I., Yagura H., Oda I. et al. Premotor cortex is involved in restoration of gait in stroke. Ann. Neurol. 2002; 52: 188-194.
43. Mori A., Waters R.S., Asanuma H. Physiological properties and patterns of projection in the cortico-cortical connections from the second somatosensory cortex to the motor cortex, area 4 gamma, in the cat. Brain Res. 1989; 504: 206-210.
44. Mulholland T.Human EEG, behavioral stillness and biofeedback. Int. J. Psychophysiol. 1995; 19(3): 263-279.
45. Nudo R.J., Milliken G.W. Reorganization of movement representations in primary motor cortex following focal ischemic infarcts in adult squirrel monkeys. J. Neurophysiol. 1996; 75: 2144-2149.
46. Pascual-Leone A., Torres F. Plasticity of the sensorimotor cortex representation of the reading finger in Braille readers. Brain 1993; 116: 39-52.
47. Pascual-Leone A., Grafman J., Hallett M. Modulation of cortical motor output maps during development of implicit and explicit knowledge. Science 1994; 263: 1287-1289.
48. Peurala S.H., Pitkanen K., Sivenius J., TarkkaI.M. How much exercise does the enhanced gait-oriented physiotherapy provide for chronic stroke patients? J. Neurol. 2004; 251: 449-453.
49. Platz T.Impairment-oriented training (IOT)-scientific concept and evidence-based treatment strategies. Restor. Neurol. Neurosci. 2004; 22: 301-315.
50. Ro T., Noser E., Boake C. et al. Functional reorganization and recovery after constraint-induced movement therapy in subacute stroke: case reports. Neurocase 2006; 12: 50-60.
51. Rosenzweig M.R. Environmental complexity, cerebral change, and behavior. Am. Psychol. 1966; 21: 321-332.
52. Seitz R.J., Azari N.P. Cerebral reorganization in man after acquired lesions. Adv. Neurol. 1999; 81: 37-47.
53. Seitz R.J., Canavan A.G., Yaguez L. et al. Successive roles of the cerebellum and premotor cortices in trajectorial learning. Neuroreport 1994; 5: 2541-2544.
54. Selivanov V., Chernikova L.A., Avdyunina I. et al. Intrapharyngeal electrostimulation in patients with post-stroke dysphagia. Neurology 2GG5; б4 (Suppl. 1): A11G.
55. Shestakova M., Lanskaya L., Chernikova L., Ioffe M. Voluntary control of EMG with or without visual feedback in healthy subjects and patients with poststroke hemiparesis. Gait & Posture 2GG5; 21 (Suppl. 1): S111.
56. Shumway-Cook A., Woollacott M.H. Motor control. Theory and practical applications. Williams & Wilkins, 1995.
57. Taub E., Miller N.E., Novack T.A. et al. Technique to improve chronic motor deficit after stroke. Arch. Phys. Med. Rehab. 1993; 74: 347-354.
58. UmarovaR.U., TanashaynM.M., ChernikovaL.A., KrotenkovaM.V. The intensity of the afferent input is the main factor for the benefit of the rehabilitation in acute stroke patients. Neurorehabilitation & Neural Repair 2GG6; 2G: 97.
59. Volpe B.T., Ferraro M., Lynch D. et al. Robotics and other devices in the treatment of patients recovering from stroke. Curr. Neurol. Neurosci. Rep. 2GG5; 5: 465-47G.
6G. Woolf C.J., SalterM.W. Neuronal plasticity: Increasing the gain in pain. Science 2GGG; 288: 17б5-17б8.
б1. You S.H., Jang S.H., Kim Y.H. et al. Virtual reality-induced cortical reorganization and associated locomotor recovery in chronic stroke. Stroke 2GG5; Зб: 11бб-1178.
Brain plasticity and modern rehabilitation technologies
L.A. Chernikova
Institute of Neurology, Russian Academy of Medical Sciences, Moscow Key words: neuroplasticity, afferent inputs, novel neurorehabilitation technologies.
Discussed are basic achievements in the studies of neuronal plasticity with the use of modern neuroimaging methods and, first of all, functional MRI. The role of various afferent inputs in these processes is emphasized. Novel neurorehabilitation technologies such as constraint-induced therapy (CI therapy), «LOCOMAT» system, the robotic therapy etc., are considered as the source of intensive goal-directed afferentation. Data concerning the possibility of the use of neuromuscular electrostimulation at the first hours after ischemic stroke are presented. The unique methods of intrapharyngeal electrostimulation in the treatment of dysphagia of different etiologies are discussed. Efficiency of transcranial electrostimulation in central post-
stroke pain syndrome is described. Data on possibilities of the EMG feedback in training of the precision grip, one of the basic motor hand skills, are presented. The details of learning of different postural tasks using postural sway feedback in patients with poststroke hemiparesis, Parkinson’s disease and spinocerebellar ataxias are discussed. Data on efficiency of alpha-stimulating training in patients with central poststroke pain syndrome and with clinical prevalence of affective disturbances are presented. Finally, prospects of one of the most interesting novel rehabilitation technologies, the technology based on virtual reality, are discussed.
Пластичность мозга и поведение
Брайан Колб, 1 Роббин Гибб и Терри Робинсон
Канадский центр поведенческой неврологии, Летбриджский университет, Летбридж, Альберта, Канада (B.K., RG.), И Департамент психологии Мичиганского университета, Анн-Арбор, Мичиган (T.R.)
Аннотация
Хотя когда-то мозг считался довольно статичным органа, теперь ясно, что организация цепи мозга постоянно изменение в зависимости от опыта.Эти изменения называются мозгом пластичность, и они связаны с функциональными изменениями, которые включают такие явления, как память, зависимость и восстановление функций. Недавнее исследование показал, что на пластичность мозга и поведение может влиять множество факторы, включая пред- и послеродовой опыт, лекарства, гормоны, созревание, старение, диета, болезни и стресс. Понимание того, как эти факторы влияние на организацию и функции мозга важно не только для понимание нормального и ненормального поведения, а также для проектирования лечение поведенческих и психологических расстройств, от зависимости до Инсульт.
Ключевые слова
зависимость; восстановление; опыт; пластичность мозга
Один из самых интригующих вопросов поведенческого нейробиология касается того, каким образом нервная система может изменять свои организации и, в конечном итоге, ее функции на протяжении всей жизни человека, свойство, которое часто называют , пластичность . Способность изменить является фундаментальной характеристикой нервной системы и проявляется даже в простейший из организмов, например крошечный червяк C.elegans , , нервная система которых имеет только 302 клетки. Когда нервная система изменяется, часто происходит коррелированное изменение поведение или психологическая функция. Это изменение поведения известно под названиями такие как обучение, память, зависимость, созревание и восстановление. Таким образом, для Например, когда люди осваивают новые двигательные навыки, например, играя в музыкальную прибора, происходят пластические изменения в структуре клеток нервной система, лежащая в основе моторики.Если пластиковые изменения как-то предотвращено возникновение моторного обучения не происходит. Несмотря на то что психологи предположили, что нервная система особенно чувствительна к опыта во время разработки, только недавно они начали оценить потенциал пластических изменений в мозге взрослого человека. Понимание пластичность мозга, очевидно, представляет значительный интерес, поскольку она обеспечивает окно для понимания развития мозга и поведения, и потому что это позволяет понять причины нормального и ненормального поведения.
ПРИРОДА ПЛАСТИЧНОСТИ МОЗГА
основное предположение исследований мозга и поведенческой пластичности состоит в том, что если изменения поведения, должны быть некоторые изменения в организации или свойствах нейронная схема, которая производит поведение. И наоборот, если нейронные сети изменились по опыту, должны быть соответствующие изменения в функциях через эти сети. Для исследователя, заинтересованного в понимании факторы, которые могут изменить мозговые цепи и, в конечном итоге, поведение, основной Задача состоит в том, чтобы найти и количественно оценить изменения.В принципе пластика меняет в нейронных цепях могут отражать либо модификации существующих схем или создание новых схем. Но как исследователи могут измерить изменения в нейронной схеме? Потому что нейронные сети состоят из отдельных нейронов, каждый из которых соединяется с подмножество других нейронов для формирования взаимосвязанных сетей, логическое место для искать пластические изменения стоит на стыках между нейронами, то есть на синапсы.Однако определить, были ли синапсы добавлены или потеряны в определенной области, учитывая, что человеческий мозг имеет что-то как 100 миллиардов нейронов, и каждый нейрон составляет в среднем несколько тысяч синапсы. Очевидно, что сканировать мозг в поисках измененных синапсов, поэтому необходимо идентифицировать и детально изучить небольшую их часть. Но какие синапсы следует изучать? Учитывая, что у нейробиологов довольно хороший представление о том, какие области мозга вовлечены в конкретное поведение, они могут сузить их поиск до вероятных областей, но по-прежнему остаются чрезвычайно сложная для изучения система.Однако существует процедура, которая облегчает работу.
В в конце 1800-х годов Камилло Гольджи изобрел технику окрашивания случайного подмножества нейронов (1-5%), так что тела клеток и дендритные деревья отдельных клетки можно визуализировать (рис. 1). Дендриты клетки функционируют как строительные леса для синапсов, так же как ветви деревьев обеспечивают место для листьев расти и подвергаться воздействию солнечного света. Полезность техники Гольджи может быть понимается, следуя этой древесной метафоре.Есть несколько способов мог оценить, сколько листьев на дереве, не считая каждый лист. Таким образом, можно было измерить общую длину ветвей дерева, а также густота листьев на представительной ветви. Затем, просто умножив Длина ветки по густоте листьев, можно оценить общую листву. Похожий процедура используется для оценки количества синапсов. Около 95% синапсов клетки находятся на его дендритах (ветвях нейрона). Кроме того, существует примерно линейная связь между пространством, доступным для синапсов (дендритные поверхность) и количество синапсов, поэтому исследователи могут предположить, что увеличивает или уменьшение дендритной поверхности отражает изменения в синаптической организации.
ФАКТОРЫ, ВЛИЯЮЩИЕ НА ПЛАСТИЧНОСТЬ МОЗГА
Используя Процедуры окрашивания по Гольджи, различные исследователи показали, что содержание животных в сложных и простых средах приводит к широким различиям в количество синапсов в определенных областях мозга. В общем, такие эксперименты показывают этот конкретный опыт украшает схему, в то время как отсутствие тех опыт не дает этого (например, Greenough & Chang, 1989). До не давнего времени, влияние этих нейропсихологических экспериментов было на удивление ограниченным, в отчасти потому, что меры по охране окружающей среды воспринимались как экстремальные и, следовательно не характерно для событий, переживаемых нормальным мозгом.Оно стало ясно, однако, не только то, что синаптическая организация изменяется с опытом, но также и то, что набор факторов, которые могут это сделать, намного шире, чем любой ожидал. Факторы, которые, как сейчас известно, влияют на структуру нейронов и поведение включает следующее:
§ опыт (как до, так и послеродовой)
§ психоактивные препараты (например, амфетамин, морфин)
§ гонадные гормоны (например,грамм., эстроген, тестостерон)
§ противовоспалительные средства (например, ингибиторы ЦОГ-2)
§ факторы роста (например, нервные фактор роста)
§ диетические факторы (например, витаминно-минеральные добавки)
§ генетические факторы (например, штамм различия, генетически модифицированные мыши)
§ болезнь (например, болезнь Паркинсона болезнь, шизофрения, эпилепсия, инсульт)
- напряжение
- мозг травмы и болезни
Мы обсуждаем два примера проиллюстрировать.
Ранний опыт
Это принято считать, что ранний опыт по-разному влияет на поведение, чем аналогичный опыт в более позднем возрасте. Причина такой разницы однако не понимается. Чтобы исследовать этот вопрос, мы поместили животных в сложная среда либо в подростковом возрасте, либо во взрослом возрасте, либо в старости (Kolb, Гибб и Горный, 2003). Мы ожидали, что будет количественные различия во влиянии опыта на синаптическую организацию, но, к нашему удивлению, мы также обнаружили качественных отличий .Таким образом, как многие исследователи до нас обнаружили, что длина дендритов и плотность синапсов была увеличена в нейронах моторной и сенсорной коры у взрослых и пожилых животных, содержащихся в сложной среде (относительно стандартная лабораторная клетка). Напротив, животные помещены в ту же среду, что и молодые особи показали увеличение длины дендритов, но уменьшение позвоночника. плотность. Другими словами, та же самая манипуляция с окружающей средой качественно различное влияние на организацию нейронных цепей у молодых особей, чем у взрослых.
Преследовать это открытие, мы позже дали детенышам 45 минут ежедневной тактильной стимуляции небольшой кисточкой (15 минут три раза в день) в течение первых 3 недель жизнь. Наши поведенческие исследования показали, что этот, казалось бы, безобидный ранний опыт улучшенные моторные и когнитивные навыки в зрелом возрасте. Анатомические исследования показали, кроме того, что у этих животных наблюдалось уменьшение позвоночника. плотность, но без изменения длины дендритов в корковых нейронах; еще один паттерн зависимых от опыта нейронных изменений.(Параллельные исследования показали другие изменения тоже, включая нейрохимические изменения, но они выходят за рамки текущее обсуждение.) Вооруженные этими результатами, мы спросили, есть ли пренатальный опыт может также изменить структуру мозга через несколько месяцев совершеннолетие. Действительно, это так. Например, потомство крысы, содержащееся в В сложной среде на сроке беременности увеличились синаптические пространство на нейронах коры головного мозга в зрелом возрасте. Хотя мы не знаем как пренатальные переживания изменяют мозг, кажется вероятным, что некоторые химические реакция матери, будь то гормональная или иная, может проникать через плаценту барьер и изменение генетических сигналов в развивающемся мозге.
Наши исследования, показывающие, что опыт может уникальным образом влиять на развивающийся мозг, привели нас к интересно, может ли травмированный мозг младенца быть восстановлен окружающей средой лечения. Мы не были удивлены, обнаружив, что посттравматический опыт, такой как тактильные поглаживания могут изменить пластичность и поведение мозга, потому что мы пришел к выводу, что такие переживания были мощными модуляторами мозга развития (Колб, Гибб и Горный, 2000). Однако удивительным было то, что этот пренатальный опыт, например, размещение беременной матери в комплексе окружающей среды, может повлиять на то, как мозг отреагирует на травму, которая получать только после рождения.Другими словами, пренатальный опыт изменил реакция мозга на травму в более позднем возрасте. Этот тип исследования имеет глубокие значение для превентивного лечения детей из группы риска по ряду неврологические расстройства.
Психоактивные препараты
Многие люди, которые принимают стимулирующие препараты, такие как никотин, амфетамин или кокаин, делают это за их мощные психоактивные эффекты. Долгосрочные поведенческие последствия злоупотребление такими психоактивными препаратами теперь хорошо задокументировано, но известно гораздо меньше о том, как повторяющееся воздействие этих препаратов влияет на нервную систему.Один экспериментальная демонстрация очень стойкой формы наркотика пластичность, зависящая от опыта, известна как поведенческая сенсибилизация. За Например, если крысе дать небольшую дозу амфетамина, она сначала покажет небольшое увеличение двигательной активности (например, движения, вставания на дыбы). Когда крыса однако при повторном приеме той же дозы активность увеличивается или сенсибилизируется, и животное может оставаться сенсибилизированным к недели, месяцы или даже годы, даже если лечение лекарствами прекращено.
Изменений в поведении, которое возникает в результате прошлого опыта и может сохраняться в течение месяцы или годы, как и воспоминания, считаются результатом изменений в моделях синаптическая организация. Параллели между лекарственной сенсибилизацией и память побудила нас спросить, сенсибилизированы ли нейроны животных к лекарствам злоупотребления демонстрируют долгосрочные изменения, аналогичные тем, которые связаны с памятью (например, Робинсон и Колб, 1999). Сравнение эффектов амфетамина и физиологический раствор на структуру нейронов в области мозга, известной как прилежащее ядро, которое опосредует психомоторные активирующие эффекты амфетамина, показали, что нейроны в мозге, обработанном амфетамином, имели больше дендритный материал, а также более плотно организованные шипы.Эти пластиковые Однако изменения не были обнаружены по всему мозгу, а скорее были локализованы в такие области, как префронтальная кора и прилежащее ядро, оба из которых считается, что они играют определенную роль в полезных свойствах этих препаратов. Потом исследования показали, что эти лекарственные изменения обнаруживаются не только при животным делают инъекции экспериментатор, но также и когда животные обучены самостоятельно принимать лекарства, что заставляет нас предполагать, что аналогичные изменения в синаптической организации можно найти у людей, наркоманов.
Прочие факторы
Все факторов, перечисленных в таблице 1, имеют эффекты, которые концептуально аналогичны к двум примерам, которые мы только что обсудили. Например, травма головного мозга нарушает синаптическая организация мозга, и когда есть функциональная улучшение после травмы, происходит коррелированная реорганизация нервной схем (например, Kolb, 1995). Но не все факторы действуют одинаково в мозг. Например, эстроген стимулирует образование синапсов в некоторых структурах. но уменьшает количество синапсов в других структурах (например,г., Колб, Форги, Гибб, Горни и Раунтри, 1998), модель изменений, которую также можно увидеть с некоторые психоактивные препараты, такие как морфин. Итак, теперь выясняется, что практически любая манипуляция, которая вызывает стойкое изменение поведения, оставляет анатомический след в головном мозге.
ВЫВОДЫ И ПРОБЛЕМЫ
Есть несколько выводов, которые можно сделать из наших исследований. Во-первых, опыт меняет мозг, и это происходит в зависимости от возраста.Во-вторых, как дородовой, так и послеродовой опыт имеют такие эффекты, и эти эффекты продолжительны и могут влияют не только на структуру мозга, но и на поведение взрослых. В-третьих, вроде бы подобный опыт может по-разному изменять нейронные цепи, хотя каждое из изменений проявляется в изменении поведения. В-четвертых, разнообразие поведенческие расстройства, от зависимости до неврологических и психиатрических расстройства коррелируют с локализованными изменениями в нервных цепях.Ну наконец то, методы лечения, направленные на изменение поведения, например, лечение зависимости, инсульт или шизофрения, вероятно, будут наиболее эффективными, если они смогут дальнейшая реорганизация соответствующих схем мозга. Кроме того, исследования нейрональных структура обеспечивает простой метод скрининга лечения, которое вероятно быть эффективным при лечении таких расстройств, как деменция. Действительно, наши исследования показывают, что новое поколение противоартритных препаратов (известных как ЦОГ-2 ингибиторы), которые уменьшают воспаление, могут обратить вспять возрастные синаптические потери и, следовательно, должны рассматриваться как полезные методы лечения возрастных когнитивная потеря.
Хотя теперь многое известно о пластичности и поведении мозга, многие теоретические вопросы оставаться. Зная, что широкий спектр опыта и агентов может изменить синаптическая организация и поведение важны, но это приводит к новому вопросу: Как это произошло? На этот вопрос нелегко ответить, и он несомненно что существует более одного ответа. Мы приводим один пример для иллюстрации.
Нейротрофический факторы — это класс химических веществ, которые, как известно, влияют на синаптические организация.Примером является фактор роста фибробластов-2 (FGF-2). Производство FGF-2 увеличивается за счет различных опытов, таких как комплексное жилье и тактильные поглаживания, а также такие наркотики, как амфетамин. Таким образом, возможно этот опыт стимулирует выработку FGF-2, а это, в свою очередь, увеличивает производство синапсов. Но опять же вопрос в том, как это сделать. Одна из гипотез состоит в том, что FGF-2 каким-то образом изменяет способ экспрессии разных генов конкретными нейронами а это, в свою очередь, влияет на то, как синапсы генерируются или теряются.В другом слова, факторы, которые изменяют поведение, в том числе опыт, могут сделать это, изменив экспрессия генов, результат, который делает традиционное соотношение генов и окружающей среды дискуссии бессмысленны.
Другое проблемы вращаются вокруг пределов и постоянства пластических изменений. После всего, люди ежедневно сталкиваются и узнают новую информацию. Есть ли предел тому, как сколько клеток может измениться? Кажется маловероятным, что клетки могут продолжать увеличиваться и добавлять синапсы на неопределенный срок, но что это контролирует? Мы видели в наших исследованиях зависимые от опыта изменения у младенцев, подростков и взрослых, которые испытывают и добавляет, и сокращает синапсы, но какие правила регулируют, когда один или может произойти другое? Этот вопрос приводит к другому, а именно: изменения в ответ на различный опыт могут взаимодействовать.Например, делает воздействие такого наркотика, как никотин, влияет на то, как мозг изменяется при обучении моторика, как игра на пианино? Рассмотрим также вопрос о постоянстве пластиковых замен. Если человек бросает курить, как долго никотин-индуцированный пластические изменения сохраняются, и влияют ли они на последующие изменения?
Один дополнительная проблема связана с ролью пластических изменений в неупорядоченном поведении. Таким образом, хотя большинство исследований пластичности предполагают, что ремоделирование нервной схема — хорошая вещь, резонно задаться вопросом, могут ли также быть основой патологического поведения.Об этой возможности известно меньше, но это кажется вероятным. Например, наркоманы часто показывают когнитивные дефицита, и кажется разумным предположить, что по крайней мере некоторые из этих дефицит может возникнуть из-за неправильной схемы, особенно в лобной доле.
В Таким образом, структура мозга постоянно меняется в ответ на неожиданно широкий спектр эмпирических факторов. Понимание того, как мозг изменения и правила, регулирующие эти изменения важны не только для понимание нормального и ненормального поведения, а также для проектирования лечение поведенческих и психологических расстройств, от зависимости до Инсульт.
Рекомендуемая литература
Колб Б. и Уишоу, I.Q. (1998). Пластичность мозга и поведение. Ежегодный обзор психологии , 49 , 43-64.
Робинсон, Т.Э., и Берридж, К.С. (в прессе). Зависимость. Ежегодный обзор психологии .
Шоу, C.A., & McEachern, J.C. (2001). К теории нейропластичности . Нью-Йорк: Тейлор и Фрэнсис.
Благодарности — Это исследование было поддержано Естественными и инженерными исследованиями. Грант Совета Б.K. и грант Национального института по борьбе со злоупотреблением наркотиками T.R ..
Примечание
1. Адресная корреспонденция Брайану Колб, CCBN, Летбриджский университет, Летбридж, Канада, T1K 3M4.
Список литературы
Greenough, W.T., & Chang, F.F. (1989). Пластичность синапса структура и структура коры головного мозга. В A. Peters & E.G. Джонс (Ред.), Кора головного мозга: Vol. 7 (стр. 391-440). Нью-Йорк: Пленум Пресс.
Колб, Б. (1995). Пластичность мозга и поведение . Махва, Нью-Джерси: Эрлбаум.
Колб Б., Форги М., Гибб Р., Горни Г. и Раунтри, С. (1998). Возраст, опыт и меняющийся мозг. Неврология и Биоповеденческие обзоры , 22 , 143-159.
Колб Б., Гибб Р. и Горни Г. (2000). Корковый пластичность и развитие поведения после ранней лобной корковой травмы. Нейропсихология развития , 18 , 423-444.
Колб Б., Гибб Р. и Горни Г. (2003). Зависящие от опыта изменения дендритной ветви и плотности шипов в неокортексе варьируются в зависимости от возраста и пола. Нейробиология обучения и памяти , X , ХХХ-ХХХ.
Робинсон Т.Э. и Колб Б. (1999). Изменения в морфология дендритов и дендритных шипов в прилежащем ядре и префронтальная кора после многократного лечения амфетамином или кокаином. Европейский Журнал неврологии , 11 , 1598-1604.
Рис. 1. Фотография нейрона. В вид слева, дендритное поле с разветвленной дендритной сетью видно. Справа — увеличенные изображения дендритных ветвей, показывающие шипы, где находится большинство синапсов. Если есть увеличение длина дендрита, плотность шипов или и то, и другое, предполагается, что их больше синапсы в нейроне.
Что такое пластичность мозга и почему она так важна?
Нейропластичность — или пластичность мозга — это способность мозга изменять свои связи или перепрограммировать себя.Без этой способности любой мозг, не только человеческий, не смог бы развиваться от младенчества до взрослой жизни или восстанавливаться после травмы мозга.
Особенность мозга заключается в том, что в отличие от компьютера он обрабатывает сенсорные и моторные сигналы параллельно. Он имеет множество нейронных путей, которые могут воспроизводить функции других, так что небольшие ошибки в развитии или временную потерю функции из-за повреждения можно легко исправить, перенаправив сигналы по другому пути.
Проблема становится серьезной, когда ошибки в развитии значительны, такие как влияние вируса Зика на развитие мозга в утробе матери или в результате повреждения в результате удара по голове или после инсульта.Тем не менее, даже в этих примерах при правильных условиях мозг может преодолеть невзгоды и восстановить некоторые функции.
Анатомия мозга гарантирует, что определенные области мозга выполняют определенные функции. Это то, что предопределено вашими генами. Например, есть область мозга, которая предназначена для движения правой руки. Повреждение этой части мозга нарушит движение правой руки. Но поскольку ощущения от руки обрабатываются другой частью мозга, вы можете чувствовать руку, но не можете ее двигать.Такое «модульное» устройство означает, что область мозга, не связанная с ощущениями или двигательной функцией, не может брать на себя новую роль. Другими словами, нейропластичность — это не синоним того, что мозг бесконечно податлив.
Частично способность организма восстанавливаться после повреждения мозга может быть объяснена улучшением состояния поврежденной области мозга, но в большинстве случаев это результат нейропластичности — образования новых нейронных связей. В исследовании Caenorhabditis elegans , типа нематоды, используемой в качестве модельного организма в исследованиях, было обнаружено, что потеря осязания усиливает обоняние.Это говорит о том, что потеря одного чувства перекладывает другие. Хорошо известно, что у людей потеря зрения в раннем возрасте может усилить другие чувства, особенно слух.
Как и у развивающегося младенца, ключом к развитию новых связей является обогащение окружающей среды, основанное на сенсорных (зрительных, слуховых, тактильных, обонятельных) и двигательных стимулах. Чем больше сенсорной и двигательной стимуляции получает человек, тем больше у него шансов выздороветь после травмы мозга. Например, некоторые из типов сенсорной стимуляции, используемые для лечения пациентов с инсультом, включают обучение в виртуальной среде, музыкальную терапию и мысленную отработку физических движений.
Основная структура мозга определяется вашими генами еще до рождения. Но его дальнейшее развитие в значительной степени зависит от процесса, называемого пластичностью развития, когда процессы развития изменяют нейроны и синаптические связи. В незрелом мозге это включает создание или потерю синапсов, миграцию нейронов через развивающийся мозг или изменение маршрута и рост нейронов.
В зрелом мозге очень мало мест, где формируются новые нейроны.Исключениями являются зубчатая извилина гиппокампа (область, связанная с памятью и эмоциями) и субвентрикулярная зона бокового желудочка, где генерируются новые нейроны, которые затем мигрируют в обонятельную луковицу (область, участвующая в обработке чувств. запаха). Хотя формирование новых нейронов таким образом не считается примером нейропластичности, оно может способствовать восстановлению мозга после повреждений.
Выращивание и обрезка
По мере роста мозга отдельные нейроны созревают, сначала отправляя несколько ветвей (аксоны, которые передают информацию от нейрона, и дендриты, которые получают информацию), а затем увеличивая количество синаптических контактов с определенными связями.
Почему не все полностью восстанавливаются после инсульта? www.shutterstock.comПри рождении каждый младенец нейрон в коре головного мозга имеет около 2500 синапсов. К двум-трем годам количество синапсов на нейрон увеличивается примерно до 15000, поскольку младенец исследует свой мир и осваивает новые навыки — процесс, называемый синаптогенезом. Но к зрелому возрасту количество синапсов уменьшается вдвое, так называемое синаптическое отсечение.
Спорный вопрос, сохраняет ли мозг способность увеличивать синаптогенез, но это может объяснить, почему агрессивное лечение после инсульта может обратить вспять повреждения, вызванные недостатком кровоснабжения определенной области мозга, путем усиления функции неповрежденных связей.
Ковка новых дорожек
Мы продолжаем иметь возможность изучать новые занятия, навыки или языки даже в пожилом возрасте. Эта сохраненная способность требует, чтобы в мозгу был доступен механизм запоминания, чтобы знания сохранялись с течением времени для последующего использования. Это еще один пример нейропластичности, который, скорее всего, связан со структурными и биохимическими изменениями на уровне синапсов.
Подкрепление или повторяющиеся действия в конечном итоге заставят мозг взрослого запомнить новое действие.По тому же механизму обогащенная и стимулирующая среда, предлагаемая поврежденному мозгу, в конечном итоге приведет к выздоровлению. Итак, если мозг настолько пластичен, почему не все, перенесшие инсульт, не могут полностью восстановить свои функции? Ответ заключается в том, что это зависит от вашего возраста (у более молодого мозга больше шансов на выздоровление), размера поврежденной области и, что более важно, лечения, предлагаемого во время реабилитации.
Пластичность мозга — ОбзорЧто такое пластичность мозга ? Означает ли это, что наш мозг из пластика? Конечно, нет.Пластичность или нейропластичность, описывает, как опыт реорганизует нервные пути в мозгу. Длинный длительные функциональные изменения в мозге происходят, когда мы узнаем что-то новое или запоминать новую информацию. Эти изменения в нейронных связях — вот что мы звоните нейропластичности .Чтобы проиллюстрировать концепцию пластичности, представьте пленку камера. Представьте, что фильм представляет ваш мозг. Теперь представьте, что используете фотоаппарат, чтобы сфотографировать дерево. Когда снимок сделан, фильм подвергается воздействию новой информации — изображения дерева.Чтобы для сохранения изображения пленка должна реагировать на свет и «изменить», чтобы записать изображение дерева. Точно так же для новых знания, которые нужно сохранить в памяти, изменения в мозгу, представляющие должны появиться новые знания. Чтобы проиллюстрировать пластичность другим способом, представьте, что производите впечатление монеты в куске глины. Для того, чтобы оттиск монеты в глине должны произойти изменения в глине — форма глина меняется, когда монета вдавливается в глину.Точно так же нейронная цепи в мозгу должны реорганизоваться в ответ на опыт или сенсорная стимуляция. Факты о нейропластичностиФАКТ 1 : Нейропластичность включает несколько различные процессы, происходящие на протяжении всей жизни.Нейропластичность не состоит из одного типа морфологическое изменение, а скорее включает несколько различных процессов, которые происходят на протяжении всей жизни человека. Многие типы клеток мозга участвует в нейропластичности, включая нейроны, глии и сосуды клетки. ФАКТ 2 : Нейропластичность имеет четкое
возрастной детерминант. Хотя пластичность происходит в течение жизни человека, разные типы пластичность преобладает в определенные периоды жизни и меньше преобладает в другие периоды. ФАКТ 3 : Нейропластичность мозга возникает в два основных условия: 1. Во время нормального развития мозга, когда сначала незрелый мозг начинает обрабатывать сенсорную информацию в зрелом возрасте (в процессе развития пластичность и пластичность обучения и памяти). 2. В качестве адаптивного механизм для компенсации утраченной функции и / или максимального увеличения оставшейся функционирует в случае травмы головного мозга. ФАКТ 4 : Окружающая среда играет ключевую роль в
влияющие на пластичность. Помимо генетических факторов, мозг формируется характеристиками окружающей человека среды и действиями той же самой человек. Пластичность развития: синаптическая обрезкаGopnick et al. (1999) описывают нейроны как растущие телефонные провода, которые общаться друг с другом.После рождения мозг новорожденного наводнен информацией из органов чувств младенца. Эта сенсорная информация должна каким-то образом вернуться в мозг, где она можно обработать. Для этого нервные клетки должны соединяться с одним другой, передающий импульсы в мозг. Продолжая аналогия с телефонным проводом, например, с протянутой основной между городами гены новорожденного указывают «путь» к правильная область мозга от конкретной нервной клетки.Например, нервные клетки сетчатки глаза посылают импульсы к первичному зрительному область в затылочной доле мозга, а не в область языка продукция (зона Вернике) в левой задней височной доле. В основные магистральные линии установлены, но конкретные соединения из одного дома в другой требуются дополнительные сигналы.В течение первых нескольких лет жизни мозг быстро растет. Поскольку каждый нейрон созревает, он посылает несколько ветвей (аксонов, которые посылают информации, и дендриты, которые принимают информацию), увеличивая количество синаптических контактов и прокладка конкретных соединений от от дома к дому, или, в случае мозга, от нейрона к нейрону.В При рождении каждый нейрон коры головного мозга имеет примерно 2500 синапсов. К тому времени, когда младенцу исполняется два или три года лет, количество синапсов составляет примерно 15000 синапсов на нейрон (Gopnick, et al., 1999). Это примерно вдвое больше, чем средний взрослый мозг. С возрастом старые связи удаляются через процесс называется synaptic pruning . Синаптическая обрезка устраняет более слабые синаптические контакты, в то время как более сильные связи сохраняются и укрепляются.Опыт определяет, какие соединения будут укреплены и обрезаны; связи которые активировались наиболее часто, сохраняются. Нейроны должны иметь цель выжить. Без цели нейроны умирают в процессе называется апоптозом, при котором нейроны, которые не получают или не передают информация становится поврежденной и умирает. Неэффективные или слабые связи «подрезать» почти так же, как садовник подрезает дерево или куст, придание растению желаемой формы. Именно пластичность позволяет процесс развития и отсечения связей, позволяющий мозгу адаптироваться себя к своей среде. Пластичность обучения и памятиКогда-то считалось, что с возрастом сети мозга становятся фиксированный. Однако за последние два десятилетия было проведено огромное количество исследований. показал, что мозг никогда не перестает изменяться и приспосабливаться. Обучение, по определению Тортора и Грабовски (1996), — это способность получить новые знания или навыки с помощью инструкций или опыта. Память — это процесс, благодаря которому это знание сохраняется с течением времени. В способность мозга изменяться с обучением — это пластичность.Так как же мозг меняется с обучением? Согласно Дурбаху (2000), появляются быть по крайней мере двумя типами модификаций, которые происходят в мозге с обучение:
Первоначально вновь полученные данные «сохраняются» в кратковременной памяти, что является временной способностью вспомнить несколько фрагментов информации.Немного данные подтверждают концепцию, что кратковременная память зависит от электрические и химические события в мозге, в отличие от структурных изменения, такие как образование новых синапсов. Одна теория краткосрочного память утверждает, что воспоминания могут быть вызваны «реверберирующими» нейронами. цепи — то есть входящий нервный импульс стимулирует первый нейрон который стимулирует второй, и так далее, ветвями от второго нейрон синапсирует с первым. Через некоторое время информация может переместиться в более постоянный тип памяти, долговременную память, которая результат анатомических или биохимических изменений, происходящих в головном мозге (Tortora, Grabowski, 1996). Пластичность, вызванная травмой: пластичность и мозг РемонтВо время восстановления мозга после травмы необходимы пластические изменения. к максимальной функциональности, несмотря на повреждение мозга. В исследованиях с участием крыс, у которых была повреждена одна область мозга, клетки мозга окружающие поврежденный участок претерпели изменения в своей функции и форма, которая позволила им взять на себя функции поврежденного клетки. Хотя этот феномен не был широко изучен на людях, данные показывают, что аналогичные (хотя и менее эффективные) изменения происходят в человеческом мозг после травмы. |
Границы | Влияние изучения пластичности мозга
Нейропластичность, также известная как нейропластичность или пластичность мозга, может быть определена как способность нервной системы изменять свою активность в ответ на внутренние или внешние раздражители путем реорганизации своей структуры, функций или связей. Фундаментальным свойством нейронов является их способность изменять силу и эффективность синаптической передачи посредством разнообразного числа зависимых от активности механизмов, обычно называемых синаптической пластичностью.Исследования прошлого века показали, что нейронная пластичность является фундаментальным свойством нервной системы у различных видов, от насекомых до человека. Действительно, исследования синаптической пластичности были не только важной движущей силой в исследованиях нейробиологии, но и вносили вклад в благополучие наших обществ, поскольку это явление связано с обучением и памятью, развитием мозга и гомеостазом, сенсорным обучением и восстановлением. от поражений головного мозга. Однако, несмотря на интенсивные исследования механизмов, управляющих синаптической пластичностью, все еще не ясно, как именно пластичность влияет на морфологию и физиологию мозга.Таким образом, изучение синаптической пластичности, несомненно, по-прежнему важно, если мы хотим полностью понять, как работает мозг.
Исторические корни концепции
Термин «пластик» происходит от латинского слова «plasticus», которое, в конечном счете, происходит от греческого термина «plastikós» или «plastos», что первоначально означало «формованный, сформированный». Однако корни современной концепции пластичности в нейробиологии еще предстоит полностью установить (Berlucchi, 2002; Stahnisch and Nitsch, 2002; Jones, 2004; DeFelipe, 2006; Berlucchi, Buchtel, 2009; Markram et al., 2011). До девятнадцатого века мозг в основном рассматривался философами (Markram et al., 2011), и, следовательно, только в конце 1800-х — начале 1900-х годов были заложены основы современной нейробиологии. В последнее десятилетие того столетия несколько ученых внесли ключевой вклад в наше современное понимание синаптической пластичности, в том числе испанский нейроанатом Сантьяго Рамон-и-Кахаль, который первым определил нейрон как анатомическую, физиологическую, генетическую и метаболическую единицу нервной системы в организме человека. его нейронная доктрина (Рамон-и-Кахал, 1899/1904; Шеперд, 1991; Джонс, 1994a).Идеи Кахаля о пластичности мозга, хотя и менее выдающиеся, сыграли важную роль в развитии концепции пластичности. В нескольких публикациях и лекциях между 1890 и 1894 годами (Ramón y Cajal, 1892, 1894a, b, c) Кахаль изложил свою гипотезу церебральной гимнастики, предположив, что способность мозга может быть увеличена за счет увеличения количества связей (Jones, 1994b ; ДеФелипе, 2006). В 1893 году итальянский нейропсихиатр Эудженио Танци предположил, что посредством особого обучения или практики повторяющаяся активность в нейронном пути может вызвать гипертрофию, тем самым укрепив уже существующие связи (Berlucchi and Buchtel, 2009).Термин «синапс» (ранее называвшийся «соединением» Кахалем) был впервые введен в употребление в 1897 году Фостером и Шеррингтоном в Кембридже, хотя в отличие от Танци они не уточняли потенциальную связь между синаптической пластичностью и обучением (Berlucchi and Buchtel, 2009; Markram et al., 2011). Позже ученик Танци Эрнесто Лугаро предположил химическую природу синаптической передачи и сформулировал связь между теориями Танци и идеями нейротропизма Кахала в 1906 и 1909 годах (Berlucchi and Buchtel, 2009).Важно отметить, что и Танци, и Лугаро были объявлены сторонниками идей Кахала (DeFelipe, 2006; Berlucchi and Buchtel, 2009). Таким образом, хотя может оставаться неясным, кто первым ввел термин «пластичность», работа Кахала, несомненно, стимулировала и повлияла на первые теории о синапсах, синаптической передаче и синаптической пластичности.
Множественные формы синаптической пластичности в двадцатом веке
В течение двадцатого века вопрос о том, как информация хранится в мозгу, стимулировал огромную работу, сосредоточенную на свойствах синаптической передачи.С публикацией в 1949 году «Организации поведения» канадский психолог Дональд Олдинг Хебб сформулировал теорию о возможных нейронных механизмах обучения и памяти (Hebb, 1949). В своей книге он провозгласил так называемые «постулаты Хебба», которые с тех пор оказали огромное влияние на исследования в области нейрофизиологии. Хотя сам Дональд Хебб признал, что он не предлагал ничего нового, и он выражает смесь веселья и раздражения, когда делается ссылка на «постулаты Хебба» (Berlucchi and Buchtel, 2009), в действительности термины «постулаты Хебба» и «пластичность Хебба» сейчас широко используются в литературе.Действительно, за год до публикации книги Хебба польский нейрофизиолог Конорски (1948) постулировал, что морфологические изменения нейронных связей могут быть субстратом обучения (Markram et al., 2011).
Первые доказательства связи краткосрочной пластичности с модификациями поведения были получены в исследованиях аплизии (Kandel and Tauc, 1965). Кратковременное облегчение и синаптическая депрессия, длящаяся от миллисекунд до минут, могут быть вызваны различными протоколами, такими как стимуляция парными импульсами или повторяющаяся высокочастотная стимуляция (Zucker and Regehr, 2002).Краткосрочная пластичность считается важной для краткосрочных реакций на сенсорные сигналы, временной модификации поведенческих состояний и краткосрочной памяти (Citri and Malenka, 2008). Спустя почти два десятилетия после появления теории Хебба, экспериментальные доказательства, подтверждающие его идеи, пришли с открытием долговременной потенциации в зубчатой извилине гиппокампа кролика (в 1975 году она называлась долговременной потенциацией или ДПН: Lømo, 2003). Эти результаты были получены благодаря ключевой технической разработке, которая происходила параллельно в лаборатории Андерсена: использованию подготовки срезов мозга (Skrede and Westgaard, 1971).Действительно, исследования с использованием препарата гиппокампальных срезов продолжали улучшать наше понимание синаптической пластичности на протяжении многих лет. Другая форма долговременной пластичности, долговременная депрессия, или LTD, была впервые предложена в 1977 году (Lynch et al., 1977). Эти достижения, наряду с разработкой внутриклеточных записей в срезах головного мозга и техник патч-зажима, привели к идентификации различных форм краткосрочной и долгосрочной пластичности в различных синапсах мозга. Например, в 80-х годах было показано, что рецепторы NMDA участвуют в синаптической пластичности (Herron et al., 1986) и постсинаптические рецепторы AMPA были важны во время LTP (Kauer et al., 1988), что является важным открытием в наших попытках понять синаптическую пластичность.
В последнее десятилетие двадцатого века важность относительного времени потенциалов действия, генерируемых пре- и постсинаптическими нейронами в моносинаптических связях, была показана при измерении в парах кортикальных нейронов (Markram et al., 1997, 2011), представляющих новая структура для изучения пластичности. Элегантность и простота этой экспериментальной парадигмы, также называемой пластичностью, зависящей от времени спайков (STDP), привлекли внимание нейробиологического сообщества.Однако дальнейшие исследования правил, управляющих STDP в разных типах нейронов и синапсов, выявили гораздо более сложный ландшафт (Markram et al., 2011). Более того, в то время была предложена другая форма стойкой синаптической пластичности, называемая метапластичностью (Abraham and Bear, 1996). Метапластичность, также известная как «пластичность синаптической пластичности», представляет собой феномен, который включает зависимые от активности изменения в функции нейронов, которые модулируют синаптическую пластичность. Роль метапластичности еще не ясна, но она может служить для поддержания синапсов в динамическом диапазоне активности, позволяя синапсам и сетям реагировать на изменяющуюся среду.В конце двадцатого века была открыта новая форма пластичности, которая действует в более длительных временных масштабах, названная гомеостатической пластичностью (Turrigiano et al., 1998; Turrigiano and Nelson, 2004). Гомеостатическая пластичность включает ряд явлений, которые уравновешивают изменения нейронной активности для поддержания гомеостаза в широком диапазоне временных и пространственных масштабов (Turrigiano, 2012). Наиболее изученный пример гомеостатической пластичности известен как синаптическое масштабирование (Turrigiano et al., 1998), которое позволяет нейронам обнаруживать изменения в их собственной скорости возбуждения с помощью набора кальций-зависимых датчиков, которые затем регулируют передачу рецепторов, тем самым увеличивая или уменьшая количество рецепторов глутамата в синаптических сайтах (Turrigiano, 2012).Взаимосвязь между STDP и гомеостатической пластичностью недостаточно изучена, и в настоящее время это интересная область исследований (Watt and Desai, 2010).
Параллельно с зависимыми от активности изменениями синаптической силы и эффективности синаптической передачи происходят структурные модификации аксонов, дендритных ветвей и морфологии шипов, явление, называемое структурной синаптической пластичностью. В частности, различные исследования коррелировали двунаправленные структурные изменения позвоночника с зависимой от активности синаптической пластичностью, т.е.е., увеличение размера позвоночника при LTP (Engert and Bonhoeffer, 1999; Matsuzaki et al., 2004) или сжатие позвоночника при LTD (Nagerl et al., 2004; Zhou et al., 2004). В настоящее время методы двухфотонной визуализации in vivo в сочетании с электрофизиологическими записями играют важную роль в выяснении взаимосвязи между функционально-структурной синаптической пластичностью и поведением. Например, формирование позвоночника наблюдалось после успешного выполнения задачи, связанной с моторной памятью (Xu et al., 2009), тогда как потеря позвоночника была связана с условием страха (Lai et al., 2012).
Проблемы и перспективы исследования пластичности
Многочисленные важные открытия механизмов LTP и LTD, такие как важность рецепторов NMDA, их временная зависимость, локус экспрессии или молекулярные механизмы, лежащие в основе LTP и LTD, вызвали интенсивные дискуссии к концу двадцатого века (Madison et al. ., 1991; Malenka, Nicoll, 1993, 1999; Malenka, Bear, 2004).Более того, открытие STDP в начале этого века вызвало интерес к влиянию времени и частоты на параметры, необходимые для индукции синаптической пластичности (Lisman and Spruston, 2005, 2010; Markram et al., 2011). Отчасти это произошло из-за того, что традиционные формы пластичности вызываются протоколами, основанными на частотах стимуляции, которые иногда далеки от физиологических, и поэтому маловероятно in vivo . С другой стороны, хотя другие формы дендритной деполяризации, а не обратный потенциал действия могут быть достаточными для индукции STDP, появляется все больше данных, показывающих, что STDP может быть вызван только одним своевременно сгенерированным единичным потенциалом действия по сравнению с EPSP (Родригес-Морено и Паулсен, 2008; Андраде-Талавера и др., 2016; Перес-Родригес и др., 2018). В самом деле, большинство правил и свойств STDP были определены in vitro , но STDP был обнаружен in vivo (Markram et al., 2011). Таким образом, ключевой задачей будущего станет определение механизмов, правил и ролей STDP in vivo (Schulz, 2010). В связи с этим будет важно определить точное влияние нейромодуляторов на STDP (Pawlak et al., 2010). Кроме того, необходимо будет разработать единую механистическую структуру, которая упрощает и объясняет огромную изменчивость свойств STDP в различных областях мозга и синапсах.Этому будет значительно способствовать четкое установление экспрессии и роли пресинаптических рецепторов NMDA (Sjöström et al., 2003; Rodríguez-Moreno and Paulsen, 2008; Abrahamsson et al., 2017; Costa et al., 2017; Bouvier et al. , 2018), особенно в свете предполагаемой метаботропной активности этих рецепторов (Nabavi et al., 2013, 2014; Dore et al., 2016, 2017). Наконец, потребуются более подробные исследования, чтобы определить недавно продемонстрированную роль глиальных клеток в синаптической пластичности (например,г., Переа и Арак, 2007; Наваррете и др., 2012; Allen, Lyons, 2018), определяя точную роль глиальных клеток в пластичности в разном возрасте.
Синаптическая пластичность является неотъемлемой частью развития и функций мозга, она важна для процессов обучения и памяти. Кроме того, временные окна для пластичности существуют во время развития, формируют связи в мозге и его активности (Hensch, 2004; Rodríguez-Moreno et al., 2013; Pérez-Rodríguez et al., 2018). Таким образом, изучение того, как возникает синаптическая пластичность и как она изменяется в течение определенных временных окон развития, предоставит ключевую информацию о том, как развивается мозг.Более того, лучшее понимание того, как синаптические модификации происходят во время обучения, памяти и / или развития, может помочь сформировать и повысить эффективность существующих протоколов на ранних этапах академического обучения. Более того, трансляционная значимость исследований синаптической пластичности на животных должна быть уточнена в будущем. Исследования тканей человека показывают, что синаптическая пластичность синапсов человека является кандидатным механизмом для обучения и памяти, хотя прямые доказательства действительного клеточного механизма отсутствуют (Mansvelder et al., 2019). Как наблюдали в исследованиях на животных, зависимые от активности синаптические изменения типа Хебба могут быть индуцированы в мозге человека in vivo , хотя и с различиями в правилах специфической пластичности (Mansvelder et al., 2019). Современные электрофизиологические методы и методы визуализации, обычно используемые в моделях на животных, могут быть использованы для экспериментов in vitro с человеческими тканями препарированных пациентов. Однако основной задачей на будущее является изучение синаптической пластичности в человеческом мозге in vivo .С этой целью неинвазивные методы, такие как транскраниальная магнитная стимуляция (ТМС), могут представлять собой шаг вперед (Polania et al., 2018).
С другой стороны, пластичность — это также явление, которое помогает восстановлению мозга после повреждений, вызванных такими событиями, как инсульт или травма. Действительно, способность манипулировать определенными нейронными путями и синапсами имеет важное значение для терапевтических и клинических вмешательств, которые улучшат наше здоровье. Многообещающие методы лечения, такие как глубокая стимуляция мозга, неинвазивная стимуляция мозга, нейрофармакология, упражнения, когнитивные тренировки или обратная связь с использованием функционального магнитного резонанса в реальном времени (Cramer et al., 2011), все основаны на нашем нынешнем понимании пластичности мозга и являются предметом интенсивных исследований различных патологий. Лучшее понимание механизмов, управляющих нейропластичностью после повреждения мозга или нервов, поможет улучшить качество жизни пациентов, в конечном итоге сэкономив национальные системы здравоохранения во всем мире. Следовательно, изучение синаптической пластичности имеет очевидные последствия, выходящие за рамки исследовательской среды. Разработчикам политики следует более подробно рассмотреть вопрос о том, как процессы обучения и памяти изменяются в процессе развития, а также о том, как мозг изменяет свою деятельность и восстанавливается после повреждений.В свете вышесказанного, такие усилия, вероятно, принесут социальные выгоды в сферах здравоохранения и образования, тем самым способствуя долгосрочному социально-экономическому планированию.
Авторские взносы
Все перечисленные авторы внесли существенный, прямой и интеллектуальный вклад в работу и одобрили ее к публикации.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Работа в нашей группе поддерживается Министерством экономики и конкуренции Испании (MINECO / FEDER: Grant BFU2015-68655-P для AR-M). PM-A поддерживается стипендиатом Хуана де ла Сьерва-Формасьона, получившего докторскую степень, от MINECO.
Список литературы
Abrahamsson, T., Chou, C.Y.C., Li, S.Y., Mancino, A., Costa, R.P., Brock, J.A., et al. (2017). Дифференциальная регуляция вызванного и спонтанного высвобождения пресинаптическими рецепторами NMDA. Neuron 96, 839–855.e835. DOI: 10.1016 / j.neuron.2017.09.030
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Андраде-Талавера, Ю., Дуке-Фериа, П., Паульсен, О., и Родригес-Морено, А. (2016). Длительная депрессия, зависящая от времени пресинаптического спайка, в гиппокампе мышей. Cereb. Cortex 26, 3637–3654. DOI: 10.1093 / cercor / bhw172
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Берлучки, Г. (2002).Происхождение термина «пластичность» в неврологии: Эрнесто Лугаро и химическая синаптическая передача. J. Hist. Neurosci. 11, 305–309. DOI: 10.1076 / jhin.11.3.305.10396
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бувье, Г., Ларсен, Р. С., Родригес-Морено, А., Паульсен, О., и Сьёстрём, П. Дж. (2018). К разрешению споров о пресинаптических рецепторах NMDA. Curr. Мнение. Neurobiol. 51, 1–7. DOI: 10.1016 / j.conb.2017.12.020
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Коста, Р.П., Мизусаки Б. Э., Шостром П. Дж. И Ван Россум М. К. (2017). Функциональные последствия пре- и постсинаптического проявления синаптической пластичности. Philos. Пер. R. Soc. Лондон. B Biol. Sci. 372: 20160153. DOI: 10.1098 / rstb.2016.0153
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Крамер, С. К., Сур, М., Добкин, Б. Х., О’Брайен, К., Сэнгер, Т. Д., Трояновский, Дж. К. и др. (2011). Использование нейропластичности для клинического применения. Мозг 134, 1591–1609.DOI: 10.1093 / brain / awr039
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Доре К., Аоу Дж. И Малинов Р. (2016). Возникновение метаботропной функции рецептора NMDA: выводы из визуализации. Фронт. Synaptic Neurosci. 8:20. DOI: 10.3389 / fnsyn.2016.00020
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Доре К., Стейн И. С., Брок Дж. А., Кастильо П. Э., Зито К. и Шостром П. Дж. (2017). Нетрадиционная передача сигналов рецептора NMDA. J. Neurosci. 37, 10800–10807. DOI: 10.1523 / JNEUROSCI.1825-17.2017
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хебб, Д. (1949). Организация поведения: нейропсихологическая теория . Нью-Йорк, штат Нью-Йорк: Wiley.
Google Scholar
Херрон К. Э., Лестер Р. А., Коан Э. Дж. И Коллингридж Г. Л. (1986). Частотно-зависимое участие рецепторов NMDA в гиппокампе: новый синаптический механизм. Природа 322, 265–268.DOI: 10.1038 / 322265a0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Kandel, E. R., and Tauc, L. (1965). Механизм гетеросинаптического облегчения в гигантской клетке брюшного ганглия Aplysia depilans . J. Physiol. 181, 28–47. DOI: 10.1113 / jphysiol.1965.sp007743
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кауэр, Дж. А., Маленка, Р. К., и Николл, Р. А. (1988). Устойчивая постсинаптическая модификация опосредует долгосрочную потенциацию в гиппокампе. Neuron 1, 911–917. DOI: 10.1016 / 0896-6273 (88)
-1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Конорски Дж. (1948). Условные рефлексы и нейронная организация . Кембридж: Издательство Кембриджского университета.
Лисман, Дж., И Спрустон, Н. (2005). Требования постсинаптической деполяризации для LTP и LTD: критика пластичности, зависящей от времени спайков. Нац. Neurosci. 8, 839–841. DOI: 10.1038 / nn0705-839
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мэдисон, Д.В., Маленка, Р. К., и Николл, Р. А. (1991). Механизмы, лежащие в основе долговременной потенциации синаптической передачи. Annu. Rev. Neurosci. 14, 379–397. DOI: 10.1146 / annurev.ne.14.030191.002115
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Malenka, R.C., и Nicoll, R.A. (1993). Зависимая от NMDA-рецепторов синаптическая пластичность: множественные формы и механизмы. Trends Neurosci. 16, 521–527. DOI: 10.1016 / 0166-2236 (93)
-TPubMed Аннотация | CrossRef Полный текст | Google Scholar
Мансвельдер, Х.Д., Верхоог М. Б., Горюнова Н. А. (2019). Синаптическая пластичность в корковых цепях человека: клеточные механизмы обучения и памяти в человеческом мозге? Curr. Мнение. Neurobiol. 54, 186–193. DOI: 10.1016 / j.conb.2018.06.013
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Markram, H., Lubke, J., Frotscher, M., and Sakmann, B. (1997). Регулирование синаптической эффективности за счет совпадения постсинаптических ПД и ВПСП. Наука 275, 213–215.DOI: 10.1126 / science.275.5297.213
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мацузаки М., Хонкура Н., Эллис-Дэвис Г. К. и Касаи Х. (2004). Структурные основы долговременной потенциации одиночных дендритных шипов. Природа 429, 761–766. DOI: 10.1038 / nature02617
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Набави, С., Фокс, Р., Альфонсо, С., Аоу, Дж., И Малинов, Р. (2014). Транспортировка GluA1 и метаботропный NMDA: рассмотрение результатов других лабораторий, несовместимых с нашими. Philos. Пер. R. Soc. Лондон. B Biol. Sci. 369: 20130145. DOI: 10.1098 / rstb.2013.0145
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Набави, С., Кессельс, Х. В., Альфонсо, С., Аоу, Дж., Фокс, Р., и Малинов, Р. (2013). Функция метаботропного рецептора NMDA необходима для длительной депрессии, зависимой от рецептора NMDA. Proc. Natl. Акад. Sci. США 110, 4027–4032. DOI: 10.1073 / pnas.1219454110
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Nagerl, U.В., Эберхорн, Н., Кембридж, С. Б., и Бонхёффер, Т. (2004). Морфологическая пластичность, зависящая от двунаправленной активности нейронов гиппокампа. Neuron 44, 759–767. DOI: 10.1016 / j.neuron.2004.11.016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Наваррете, М., Переа, Г., Фернандес Де Севилья, Д., Гомес-Гонсало, М., Нуньес, А., Мартин, Э. Д. и др. (2012). Астроциты обеспечивают in vivo холинергических синаптических пластичностей. PLoS Biol. 10: e1001259. DOI: 10.1371 / journal.pbio.1001259
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Павлак В., Виккенс Дж. Р., Кирквуд А. и Керр Дж. Н. (2010). Время — это еще не все: нейромодуляция открывает ворота STDP. Фронт. Synaptic Neurosci. 2: 146. DOI: 10.3389 / fnsyn.2010.00146
CrossRef Полный текст | Google Scholar
Перес-Родригес, М., Арройо-Гарсия, Л. Э., Приус-Менгуаль, Дж., Андраде-Талавера, Ю., Арменгол, Дж.А., Перес-Вильегас, Э. М. и др. (2018). Опосредованная аденозиновым рецептором потеря развития спайк-зависимой депрессии в гиппокампе. Cereb. Cortex . DOI: 10,1093 / cercor / bhy194. [Epub перед печатью].
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Поляния Р., Ницше М. А. и Рафф К. С. (2018). Изучение и изменение функции мозга с помощью неинвазивной стимуляции мозга . Nat. Neurosci. 21, 174–187. DOI: 10.1038 / s41593-017-0054-4
CrossRef Полный текст
Рамон-и-Кахаль, С.(1892). El nuevo Concepto de la histología en los centros nerviosos. Rev. Ciencias Méd. Барселона 18, 361–376, 457–476, 505–520, 529–541.
Google Scholar
Рамон-и-Кахаль, С. (1894a). Обсуждаем генеральные морфологи í а-ля Célula Nerviosa . Мадрид: Моя.
Рамон-и-Кахаль, С. (1894b). Обсуждаем родовые морфологии нервной целюлы. La Veterinaria Española 37, 257–260, 273–275, 289–291.
Google Scholar
Рамон-и-Кахаль, С. (1894c). Лекция кроона: тонкая структура нервных центров. Proc. Royal Soc. Лондон. 55, 444–468. DOI: 10.1098 / rspl.1894.0063
CrossRef Полный текст | Google Scholar
Рамон-и-Кахаль, С. (1899/1904). Textura del Sistema Nervioso del Hombre y de los Vertebrados . Мадрид: Моя.
Родригес-Морено, А., Гонсалес-Руэда, А., Банерджи, А., Аптон, А. Л., Крейг, М. Т., и Паулсен, О.(2013). Пресинаптическая самодепрессия при развитии неокортикальных синапсов. Neuron 77, 35–42. DOI: 10.1016 / j.neuron.2012.10.035
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шеперд, Г. М. (1991). Основы нейронной доктрины . Нью-Йорк, Нью-Йорк: Издательство Оксфордского университета.
Google Scholar
Сьёстрём П. Дж., Турриджано Г. Г. и Нельсон С. Б. (2003). Neocortical LTD через одновременную активацию пресинаптических NMDA и каннабиноидных рецепторов. Neuron 39, 641–654. DOI: 10.1016 / S0896-6273 (03) 00476-8
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Скреде К. К. и Вестгаард Р. Х. (1971). Поперечный срез гиппокампа: хорошо выраженная кортикальная структура, поддерживаемая in vitro . Brain Res. 35, 589–593. DOI: 10.1016 / 0006-8993 (71)
-7PubMed Аннотация | CrossRef Полный текст | Google Scholar
Stahnisch, F. W., and Nitsch, R. (2002). Концепция нейрональной пластичности Сантьяго Рамона и Кахала: неоднозначность продолжает жить. Trends Neurosci. 25, 589–591. DOI: 10.1016 / S0166-2236 (02) 02251-8
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Турриджиано, Г. (2012). Гомеостатическая синаптическая пластичность: локальные и глобальные механизмы стабилизации нейрональной функции. Колд Спринг Харб. Перспектива. Биол. 4: a005736. DOI: 10.1101 / cshperspect.a005736
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Турриджиано, Г. Г., Лесли, К. Р., Десаи, Н.С., Резерфорд, Л. С., и Нельсон, С. Б. (1998). Масштабирование квантовой амплитуды в нейронах неокортекса в зависимости от активности. Природа 391, 892–896. DOI: 10.1038 / 36103
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Xu, T., Yu, X., Perlik, A.J., Tobin, W.F., Zweig, J.A., Tennant, K., et al. (2009). Быстрое формирование и избирательная стабилизация синапсов для устойчивых моторных воспоминаний. Природа 462, 915–919. DOI: 10.1038 / nature08389
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чжоу, Q., Хомма, К. Дж., И Пу, М. М. (2004). Сморщивание дендритных шипов, связанное с длительной депрессией синапсов гиппокампа. Neuron 44, 749–757. DOI: 10.1016 / j.neuron.2004.11.011
PubMed Аннотация | CrossRef Полный текст | Google Scholar
ученых MIT открыли фундаментальное правило пластичности мозга | MIT News
Наш мозг известен своей гибкостью, или «пластичностью», потому что нейроны могут делать новые вещи, создавая новые или более сильные связи с другими нейронами.Но если некоторые связи укрепляются, рассуждают нейробиологи, нейроны должны компенсировать это, чтобы не перегружать их входными данными. В новом исследовании, опубликованном в журнале Science , исследователи из Института обучения и памяти Пикауэра при Массачусетском технологическом институте впервые демонстрируют, как достигается этот баланс: когда одно соединение, называемое синапсом, усиливается, сразу же соседние синапсы ослабевают под действием важнейший белок под названием Arc.
Старший автор Мриганка Сур сказал, что был взволнован, но не удивлен, что его команда обнаружила простое, фундаментальное правило, лежащее в основе такой сложной системы, как мозг, где у 100 миллиардов нейронов есть тысячи постоянно меняющихся синапсов.Он сравнивает это с тем, как огромная коса рыб может внезапно менять направление в массе, пока ведущая рыба поворачивается, а все остальные рыбы подчиняются простому правилу — следовать за рыбой прямо перед ней.
«Коллективное поведение сложных систем всегда подчиняется простым правилам, — говорит Сур, профессор нейробиологии Пола Э. и Лайлы Ньютон в Институте Пикауэра и на кафедре мозговых и когнитивных наук Массачусетского технологического института. «Когда один синапс поднимается вверх, в пределах 50 микрометров происходит снижение прочности других синапсов с использованием четко определенного молекулярного механизма.”
Это открытие, по его словам, дает объяснение того, как синаптическое усиление и ослабление сочетаются в нейронах, создавая пластичность.
Множественные манипуляции
Хотя правило, которое они нашли, было простым, эксперименты, которые его выявили, не подтвердили. Ведущие авторы Сами Эль-Бустани и Жак Пак Кан Ип, постдоки в лаборатории Сура, работали над активацией пластичности в зрительной коре головного мозга мышей, а затем отслеживали, как изменились синапсы, чтобы это произошло.
В одном из ключевых экспериментов они задействовали пластичность, изменив «рецептивное поле» нейрона или участок поля зрения, на которое он реагирует. Нейроны получают входные данные через синапсы на маленьких шипах своих ветвистых дендритов. Чтобы изменить рецептивное поле нейрона, ученые точно определили шип на соответствующем дендрите нейрона, а затем внимательно наблюдали за изменениями в его синапсах, когда они показывали мыши цель в определенном месте на экране, которая отличалась от исходной рецептивной способности нейрона. поле.Всякий раз, когда цель оказывалась в новом положении рецептивного поля, которое они хотели вызвать, они усиливали реакцию нейрона, мигая синим светом внутри зрительной коры головного мозга мыши, вызывая дополнительную активность, как это может сделать другой нейрон. Нейрон был генетически сконструирован таким образом, чтобы его можно было активировать световыми вспышками — метод, названный «оптогенетикой».
Исследователи делали это снова и снова. Поскольку световая стимуляция коррелировала с каждым появлением цели в новом положении в поле зрения мыши, это заставляло нейрон усиливать определенный синапс на позвоночнике, кодируя новое рецептивное поле.
«Я думаю, это удивительно, что мы можем перепрограммировать отдельные нейроны в неповрежденном мозге и наблюдать в живой ткани разнообразие молекулярных механизмов, которые позволяют этим клеткам интегрировать новые функции за счет синаптической пластичности», — говорит Эль-Бустани.
По мере того как синапс для нового рецептивного поля рос, исследователи могли видеть под двухфотонным микроскопом, что близлежащие синапсы также уменьшились. Они не наблюдали этих изменений в экспериментальных контрольных нейронах, лишенных оптогенетической стимуляции.
Но затем они пошли дальше, чтобы подтвердить свои выводы. Поскольку синапсы такие крошечные, они близки к пределу разрешения световой микроскопии. Поэтому после экспериментов команда исследовала ткани мозга, содержащие дендриты управляемых и управляемых нейронов, и отправила их соавторам в Федеральную политехническую школу Лозанны в Швейцарии. Они выполнили специальную трехмерную электронную микроскопию с более высоким разрешением, подтвердив, что структурные различия, наблюдаемые под двухфотонным микроскопом, действительны.
«Это самая длинная длина дендрита, когда-либо реконструированная после визуализации in vivo», — сказал Сур, который также руководит Центром Саймонса для социального мозга в Массачусетском технологическом институте.
Конечно, перепрограммирование генно-инженерного нейрона мыши с помощью вспышек света — неестественная манипуляция, поэтому команда провела еще один, более классический эксперимент по «монокулярной депривации», в котором они временно закрыли один глаз мыши. Когда это происходит, синапсы в нейронах, связанных с закрытым глазом, ослабевают, а синапсы, связанные с еще открытым глазом, усиливаются.Затем, когда они снова открыли ранее закрытый глаз, синапсы снова перестраиваются. Они также отслеживали это действие и видели, что по мере усиления синапсов их непосредственные соседи будут ослабевать, чтобы компенсировать это.
Раскрытие тайны Арки
Увидев новое правило в действии, исследователи все еще стремились понять, как нейроны подчиняются ему. Они использовали химическую метку, чтобы наблюдать, как ключевые рецепторы «AMPA» изменяются в синапсах, и увидели, что увеличение и усиление синапсов коррелирует с большей экспрессией рецептора AMPA, а сокращение и ослабление коррелирует с меньшей экспрессией рецептора AMPA.
Белок Arc регулирует экспрессию рецептора AMPA, поэтому команда поняла, что им нужно отслеживать Arc, чтобы полностью понять, что происходит. Проблема, по словам Сура, в том, что никто никогда раньше не делал этого в мозгу живого, ведущего себя животного. Поэтому команда обратилась к соавторам из Высшей школы медицины Киотского университета и Токийского университета, которые изобрели химическую метку, которая могла бы это сделать.
Используя тег, команда смогла увидеть, что укрепляющие синапсы были окружены ослабленными синапсами, которые увеличили экспрессию Arc.Синапсы с уменьшенным количеством Arc были способны экспрессировать больше рецепторов AMPA, тогда как увеличение Arc в соседних шипах заставляло эти синапсы экспрессировать меньше рецепторов AMPA.
«Мы думаем, что Arc поддерживает баланс синаптических ресурсов», — говорит Ип. «Если что-то идет вверх, что-то должно падать. Это главная роль Арк ».
Сур говорит, что исследование, таким образом, решает загадку Arc: никто раньше не понимал, почему Arc, по-видимому, активируется в дендритах, претерпевающих синаптическую пластичность, даже несмотря на то, что это ослабляет синапсы, но теперь ответ был ясен.Усиление синапсов увеличивает Arc, чтобы ослабить своих соседей.
Сур добавил, что это правило помогает объяснить, как обучение и память могут работать на уровне отдельного нейрона, поскольку оно показывает, как нейрон приспосабливается к повторяющейся симуляции другого нейрона.
Аня Маевска, доцент кафедры нейробиологии Центра визуальных наук Университета Рочестера, говорит, что передовые методы исследования позволили команде достичь важных новых результатов.
«Из-за сложности мониторинга и манипулирования крошечными и многочисленными синапсами, которые соединяют нейроны, большинство исследований проводилось в сокращенных препаратах с искусственными стимулами, из-за чего неясно, как идентифицированные механизмы на самом деле реализуются в сложных цепях, которые функционируют внутри мозга. реагируя на окружающую среду », — говорит Маевска.«Это новое исследование лаборатории Sur имеет огромное влияние, поскольку оно сочетает в себе передовые методы визуализации и генетические инструменты для прекрасного мониторинга функции отдельных синапсов внутри мозга, который реагирует на поведенческие стимулы, вызывающие изменения в ответах нейронов.
«Учитывая результаты этого подхода Tour de Force, мы можем теперь сказать, что в неповрежденном мозге синапсы, которые находятся в непосредственной близости друг от друга, взаимодействуют во время изменений в функции цепи через механизм, который включает молекулярный каскад, в котором играет дуга важная роль », — сказала она.«Эта информация позволяет нам понять не только то, как нейронные цепи развиваются и модифицируются в физиологических условиях, но и дает ключи, которые будут важны для определения того, как эти процессы идут наперекосяк при различных неврологических заболеваниях».
Помимо Сура, Эль-Бустани и Ипа, другими авторами статьи являются Винсент Бретон-Провенчер, Грэхам Нотт, Хироюки Окуно и Харухико Бито.
Финансирование исследований поступило от Инновационного фонда института Пикауэра, Центра Саймонса для социального мозга, стипендии Марии Кюри для получения докторской степени, долгосрочной стипендии программы Human Frontier Science Programme, Национальных институтов здравоохранения, Национального научного фонда и KAKENHI. .
Пластичность мозга — обзор
B Гипотеза синаптического гомеостаза (SHY)
Одной из наиболее влиятельных гипотез о функции сна в отношении пластичности мозга является гипотеза синаптического гомеостаза (SHY), впервые описанная Тонони и Сирелли (Тонони И Cirelli, 2003). SHY предполагает универсальную, основополагающую роль сна для пластичности мозга и когнитивных функций, восстанавливая синаптический гомеостаз. Согласно SHY, нейронная сеть отягощена зависимым от использования LTP-подобным увеличением синаптической силы (вызванным, главным образом, синаптической потенциацией) в период бодрствования и синаптогенезом во время развития.Синаптическая потенциация, в свою очередь, отражается в гомеостатической регуляции медленноволновой активности, которая увеличивается со временем нахождения в бодрствовании. Во время последующего сна происходит глобальное синаптическое даунскейлинг (рис. 28.1, верхний ряд, середина справа), что связано с постепенным снижением медленноволновой активности в процессе сна. Эта глобальная нормализация синаптических сил объясняет благотворное влияние сна на производительность и на приобретение, консолидацию и интеграцию памяти (Tononi & Cirelli, 2003, 2014).
В самом деле, многочисленные исследования на молекулярном, электрофизиологическом и структурном уровнях предоставляют доказательства того, что чистая синаптическая сила претерпевает изменения, зависящие от сна и бодрствования. В частности, экспериментальные результаты демонстрируют чистое синаптическое увеличение, преобладающее во время бодрствования, и глобальное синаптическое уменьшение масштаба, происходящее во время сна (Bushey, Tononi, & Cirelli, 2011; De Vivo et al., 2017; Diering et al., 2017; Gilestro, Tononi, & Cirelli, 2009; Huber et al., 2012; Kuhn et al., 2016; Maret, Faraguna, Nelson, Cirelli, & Tononi, 2011; Norimoto et al., 2018; Вязовский, Чирелли, Пфистер-Генскоу, Фарагуна и Тонони, 2008). Согласно SHY, во время бодрствования обучение и постоянное взаимодействие с окружающей средой запускают пластические изменения в мозге и, соответственно, увеличивают синаптическую силу, в основном за счет синаптического потенцирования и образования новых синапсов. Однако такое непрерывное увеличение синаптической силы обременяет нейрон как энергетически (например, менее «экономичное» возбуждение), так и информационным (например, снижение избирательности).Биологические затраты — это повышенное потребление энергии и более высокие потребности в материалах, поддерживающих клеточные функции, что в конечном итоге приводит к высокому клеточному стрессу. Более того, кодирование накапливаемой информации снижает отношение сигнал / шум, что отражается в (i) расширении нейронного распределения входных паттернов; (ii) пониженная селективность; и, как следствие, (iii) насыщенная способность к обучению. Соответственно, нейронам в какой-то момент необходимо перенормировать чистую синаптическую силу и восстановить синаптический гомеостаз (Tononi & Cirelli, 2014).
SHY предполагает, что эта глобальная реорганизация происходит во время сна, особенно во время сна с медленной волной, когда мозг отключен от окружающей среды и не подвержен (сознательным) внешним взаимодействиям (Massimini et al., 2005). Спонтанная «автономная» активность во время глубокого сна позволяет нейронам «всесторонне» собирать и интегрировать всю информацию, полученную в более широком временном масштабе, от эволюционного наследия до развития и острых взаимодействий с окружающей средой и обучения (Edelman, 1987; Tononi, Edelman, & Sporns, 1998).Согласно SHY, синаптическая очистка происходит во время медленноволнового сна за счет зависимой от активности синаптической депрессии или даунскейлинга. Другими словами, глобальная синаптическая сила пропорционально снижается во время сна, синапсы, находящиеся ниже минимального порогового значения силы, становятся практически неэффективными, что способствует поддержанию утилитарной сетевой коммуникации и подавлению «дорогих» избыточных синаптических связей. Таким образом, клеточные функции восстанавливаются, а нейронная сеть «перекалибрована», поддерживая функциональную селективность нейронов и улучшая соотношение сигнал / шум.Следовательно, способность кодировать новую информацию за счет ассоциативной пластичности увеличивается, а задачи и умственные способности улучшаются.
Существуют многочисленные молекулярные, электрофизиологические и структурные доказательства того, что синаптическое масштабирование происходит во время сна (Hinard et al., 2012; Huber et al., 2008; Huber, Tononi, & Cirelli, 2007; Lubenov & Siapas, 2008; Maret et al. ., 2011). На молекулярном уровне, например, экспрессия или удаление AMPArs в синаптической мембране отражает основные механические изменения, опосредующие синаптическую потенциацию и депрессию, соответственно (Kessels & Malinow, 2009).Недавнее исследование показало, что во время сна низкие уровни нейромодулятора норадреналина позволяют экспрессировать непосредственный ранний ген Homer1a в возбуждающих синаптических участках, вызывая ослабление синапсов за счет удаления или дефосфорилирования AMPAr (Diering et al., 2017). Другое недавнее исследование продемонстрировало зависимое от сна сокращение интерфейса синаптический аксон-позвоночник (ASI), предоставив дополнительные структурные доказательства того, что сон способствует уменьшению размера синапсов (De Vivo et al., 2017). На электрофизиологическом уровне синаптическая сила отражается в наклоне вызванных ответов, вызванных электрической стимуляцией у животных (Вязовский и др., 2008) или ТМС у человека (Huber et al., 2012). Исследования на животных и людях показали, что снижение наклона корковых реакций коррелирует со снижением медленноволновой активности, предположительно отражая перенормировку синаптических сил. Интересно, что результаты недавнего исследования подтверждают медленноволновую зависимость подавления синаптической регуляции за счет ингибирования резких волн гиппокампа (т. Е. Кратковременных возбуждающих всплесков («пульсационных» колебаний 200 Гц), возникающих в гиппокампе) во время медленноволнового сна, которые в Turn перехватил синаптическое даунскейлинг и нарушил приобретение новых воспоминаний (Norimoto et al., 2018). Короче говоря, основное утверждение SHY заключается в том, что сон выполняет важную универсальную функцию. Эта функция заключается в сохранении синаптического гомеостаза, которому препятствуют постоянные пластические изменения, происходящие во время бодрствования, в основном смещенные в сторону чистого увеличения синаптической силы, которое является результатом массивного синаптогенеза во время развития и обучения во взрослом возрасте (Tononi & Cirelli, 2012).
Нейропластичность в стареющем мозге
Броди, Х. Организация коры головного мозга.III. Исследование старения в коре головного мозга человека. J. Comp. Neurol. 102 , 511–516 (1955).
CAS PubMed Google ученый
Коулман, П. Д. и Флуд, Д. Г. Число нейронов и протяженность дендритов при нормальном старении и болезни Альцгеймера. Neurobiol. Старение 8 , 521–545 (1987).
CAS PubMed Google ученый
Болл, М.J. Потеря нейронов, нейрофибриллярные сплетения и грануловакуолярная дегенерация в гиппокампе при старении и деменции. Количественное исследование. Acta Neuropathol. (Berl.) 37 , 111–118 (1977).
CAS Google ученый
Бриззи, К. Р., Орди, Дж. М. и Бартус, Р. Т. Локализация клеточных изменений в мультимодальных сенсорных областях в мозге старых обезьян: возможные последствия для когнитивной потери, связанной с возрастом. Neurobiol. Старение 1 , 45–52 (1980).
CAS PubMed Google ученый
Моррисон, Дж. Х. и Хоф, П. Р. Жизнь и смерть нейронов в стареющем мозге. Наука 278 , 412–419 (1997).
CAS PubMed Google ученый
Вест, М. Дж. Новые стереологические методы подсчета нейронов. Neurobiol.Старение 14 , 275–285 (1993).
CAS PubMed Google ученый
Паккенберг, Б. и Гундерсен, Х. Дж. Число нейронов неокортекса у людей: влияние пола и возраста. J. Comp. Neurol. 384 , 312–320 (1997).
CAS PubMed Google ученый
Вест, М. Дж., Коулман, П. Д., Флад, Д. Г. и Тронкосо, Дж.C. Различия в характере потери нейронов гиппокампа при нормальном старении и болезни Альцгеймера. Ланцет 344 , 769–772 (1994). Стереологически контролируемое исследование, сообщающее о сохранении количества нейронов в большинстве субрегионов гиппокампа здоровых пожилых людей, в отличие от людей с болезнью Альцгеймера, у которых наблюдается значительное снижение количества клеток.
CAS PubMed Google ученый
Меррилл, Д.А., Робертс, Дж. А. и Тушински, М. Х. Сохранение числа и размеров нейронов в энторинальных слоях коры II, III и V / VI старых приматов. J. Comp. Neurol. 422 , 396–401 (2000).
CAS PubMed Google ученый
Петерс, А., Лиху, Д., Мосс, М. Б. и МакНалли, К. Дж. Влияние старения на область 46 лобной коры головного мозга макаки-резуса. Cereb. Cortex 4 , 621–635 (1994).
CAS PubMed Google ученый
Газзалей А. Х., Таккер М. М., Хоф П. Р. и Моррисон Дж. Х. Сохраненное количество нейронов энторинального слоя коры II у старых макак. Neurobiol. Старение 18 , 549–553 (1997).
CAS PubMed Google ученый
Кекер, Дж. И., Луйтен, П. Г. и Фукс, Э. Сохранение числа нейронов гиппокампа у старых макак-резусов. Neurobiol. Старение 24 , 157–165 (2003).
PubMed Google ученый
Меррилл Д. А., Чиба А. А. и Тушински М. Х. Сохранение количества и размера нейронов в энторинальной коре у пожилых крыс с поведенческими характеристиками. J. Comp. Neurol. 438 , 445–456 (2001).
CAS PubMed Google ученый
Рапп, П.Р. и Галлахер, М. Количество сохранившихся нейронов в гиппокампе старых крыс с дефицитом пространственного обучения. Proc. Natl Acad. Sci. USA 93 , 9926–9930 (1996). Подобно результатам, полученным на людях, этот стереологически контролируемый эксперимент показывает, что у старых крыс нет потери нейронов, и поэтому гибель клеток не может объяснять возрастные нарушения поведения.
CAS PubMed Google ученый
Расмуссен, Т., Schliemann, T., Sorensen, J. C., Zimmer, J. & West, M. J. У старых крыс с нарушением памяти: нет потери основных нейронов гиппокампа и субикулярных нейронов. Neurobiol. Старение 17 , 143–147 (1996).
CAS PubMed Google ученый
Смит, Д. Э., Рапп, П. Р., Маккей, Х. М., Робертс, Дж. А. и Тушински, М. Х. Нарушение памяти у старых приматов связано с очаговой гибелью корковых нейронов и атрофией подкорковых нейронов. J. Neurosci. 24 , 4373–4381 (2004).
CAS PubMed Google ученый
Шейбель М. Э., Линдси Р. Д., Томиясу У. и Шейбель А. Б. Прогрессивные дендритные изменения в стареющей лимбической системе человека. Exp. Neurol. 53 , 420–430 (1976).
CAS PubMed Google ученый
Шейбель, А.Б. Гиппокамп: организационные модели здоровья и старения. мех. Aging Dev. 9 , 89–102 (1979).
CAS PubMed Google ученый
Buell, S. J. и Coleman, P. D. Количественные доказательства избирательного роста дендритов при нормальном старении человека, но не при старческой деменции. Brain Res. 214 , 23–41 (1981).
CAS PubMed Google ученый
Buell, S.Дж. И Коулман, П. Д. Рост дендритов в мозге пожилого человека и нарушение роста при старческом слабоумие. Наука 206 , 854–856 (1979).
CAS PubMed Google ученый
Флад, Д. Г., Бьюэлл, С. Дж., Дефиоре, К. Х., Хорвиц, Г. Дж. И Коулман, П. Д. За возрастным ростом дендритов в зубчатой извилине человеческого мозга следует регресс у «самых старых». Brain Res. 345 , 366–368 (1985).
CAS PubMed Google ученый
Флад, Д. Г., Буэлл, С. Дж., Хорвиц, Г. Дж. И Колеман, П. Д. Дендритная степень в гранулярных клетках зубчатой извилины человека при нормальном старении и старческой деменции. Brain Res. 402 , 205–216 (1987).
CAS PubMed Google ученый
Хэнкс, С. Д. и Флуд, Д. Г. Региональная стабильность дендритной протяженности при нормальном старении человека и регресс при болезни Альцгеймера.I. CA1 гиппокампа. Brain Res. 540 , 63–82 (1991).
CAS PubMed Google ученый
Флад, Д. Г., Гуарначча, М. и Колман, П. Д. Дендритная степень в пирамидных нейронах СА2–3 гиппокампа человека при нормальном старении и старческой деменции. Brain Res. 409 , 88–96 (1987).
CAS PubMed Google ученый
Флуд, Д.G. Регионально-специфическая стабильность дендритной протяженности при нормальном старении человека и регресс при болезни Альцгеймера. II. Субикулюм. Brain Res. 540 , 83–95 (1991).
CAS PubMed Google ученый
Флад Д.Г. Критические вопросы анализа степени дендрита у стареющих людей, приматов и грызунов. Neurobiol. Старение 14 , 649–654 (1993).
CAS PubMed Google ученый
Тернер Д.A. & Deupree, D. L. Функциональное удлинение нейронов гиппокампа CA1 с возрастом у крыс Fischer 344. Neurobiol. Старение 12 , 201–210 (1991).
CAS PubMed Google ученый
Пьяпали, Г. К. и Тернер, Д. А. Повышенная протяженность дендритов в нейронах СА1 гиппокампа у старых крыс F344. Neurobiol. Старение 17 , 601–611 (1996).
CAS PubMed Google ученый
Маркхэм, Дж.А., Маккиан, К. П., Строуп, Т. С. и Джураска, Дж. М. Половое диморфное старение дендритной морфологии в СА1 гиппокампа. Гиппокамп 15 , 97–103 (2005).
CAS PubMed Google ученый
Гриль Д. и Риддл Д. Р. Возрастные и ламинарно-специфические дендритные изменения в медиальной лобной коре головного мозга крысы. Brain Res. 937 , 8–21 (2002).
CAS PubMed Google ученый
Маркхэм, Дж.А. и Джураска, Дж. М. Старение и пол влияют на анатомию передней поясной коры крыс. Neurobiol. Старение 23 , 579–588 (2002).
PubMed Google ученый
де Брабандер, Дж. М., Крамерс, Р. Дж. И Уйлингс, Х. Б. Слоистая дендритная регрессия пирамидных клеток со старением в префронтальной коре головного мозга человека. евро. J. Neurosci. 10 , 1261–1269 (1998).
CAS PubMed Google ученый
Юлингс, Х.Б. и де Брабандер, Дж. М. Нейрональные изменения при нормальном старении человека и болезнь Альцгеймера. Brain Cogn. 49 , 268–276 (2002).
PubMed Google ученый
Williams, R. S. & Matthysse, S. Возрастные изменения в мозге с синдромом Дауна и клеточная патология болезни Альцгеймера. Прог. Brain Res. 70 , 49–67 (1986).
CAS PubMed Google ученый
Курчо, К.А. и Хайндс, Дж. У. Стабильность синаптической плотности и объема шипов в зубчатой извилине у старых крыс. Neurobiol. Старение 4 , 77–87 (1983).
CAS PubMed Google ученый
Uemura, E. Возрастные изменения субикулюма Macaca mulatta : синаптическая плотность. Exp. Neurol. 87 , 403–411 (1985).
CAS PubMed Google ученый
Барнс, К.A. Нормальное старение: регионально-специфические изменения синаптической передачи в гиппокампе. Trends Neurosci. 17 , 13–18 (1994). Исчерпывающий обзор изменений в синаптической передаче, специфичных для области гиппокампа, наряду с функциональной сохранностью, который бросил вызов традиционной концепции старения как процесса общего ухудшения.
CAS PubMed Google ученый
Барнс, К. А. Дефицит памяти, связанный со старением: нейрофизиологическое и поведенческое исследование на крысах. J. Comp. Physiol. Psychol. 93 , 74–104 (1979). Первый отчет у старых крыс об увеличенной скорости распада LTP в синапсе перфорантного пути и гранулярных клеток, который коррелирует со скоростью забывания пространственной проблемы в задаче круговой платформы Барнса.
CAS PubMed Google ученый
Барнс, К. А., Рао, Г., Фостер, Т. К. и Макнотон, Б. Л. Региональные возрастные эффекты на чувствительность AMPA: электрофизиологические доказательства потери синаптических контактов в поле СА1 гиппокампа. Гиппокамп 2 , 457–468 (1992).
CAS PubMed Google ученый
Сигал, М. Изменения в действиях нейротрансмиттеров в гиппокампе старых крыс. Neurobiol. Старение 3 , 121–124 (1982).
CAS PubMed Google ученый
Лэндфилд, П. В. и Питлер, Т. А. Длительные Ca 2+ -зависимые постгиперполяризации в нейронах гиппокампа старых крыс. Наука 226 , 1089–1092 (1984). Первое сообщение о значительном увеличении K + -зависимой постгиперполяризации старых пирамидных клеток гиппокампа CA1, которая блокируется низкими концентрациями Ca 2+ .
CAS PubMed Google ученый
Niesen, C.E., Baskys, A. & Carlen, P.L. Обратное действие этанола на проводимость калия в нейронах зубчатых гранул старого гиппокампа. Brain Res. 445 , 137–141 (1988).
CAS PubMed Google ученый
Керр, Д. С., Кэмпбелл, Л. В., Хао, С. Ю. и Лэндфилд, П. В. Кортикостероидная модуляция потенциалов гиппокампа: усиление эффекта с возрастом. Наука 245 , 1505–1509 (1989).
CAS PubMed Google ученый
Потье, Б., Ламур, Ю. и Дутар, П. Возрастные изменения свойств пирамидных нейронов гиппокампа у линий крыс. Neurobiol. Старение 14 , 17–25 (1993).
CAS PubMed Google ученый
Potier, B., Rascol, O., Jazat, F., Lamour, Y. & Dutar, P. Изменения свойств пирамидных нейронов гиппокампа у старых крыс. Неврология 48 , 793–806 (1992).
CAS PubMed Google ученый
Питлер Т. и Лэндфилд П. У. Связанное со старением удлинение продолжительности кальциевого выброса в нейронах гиппокампа крысы. Brain Res. 508 , 1–6 (1990).
CAS PubMed Google ученый
Барнс, К. А. и Макнотон, Б. Л. Физиологическая компенсация потери афферентных синапсов в гранулярных клетках гиппокампа крысы во время старения. J. Physiol. (Лондон) 309 , 473–485 (1980).
CAS Google ученый
Рейнольдс, Дж. Н. и Карлен, П. Л. Снижение кальциевых потоков в старых нейронах гранул зубчатой извилины гиппокампа. Brain Res. 479 , 384–390 (1989).
CAS PubMed Google ученый
Luebke, J. I. & Rosene, D. L.Старение изменяет морфологию дендритов, входное сопротивление и подавляющую передачу сигналов в зубчатых гранулярных клетках макаки-резуса. J. Comp. Neurol. 460 , 573–584 (2003).
PubMed Google ученый
Мойер, Дж. Р. Младший, Томпсон, Л. Т., Блэк, Дж. П. и Дистерхофт, Дж. Ф. Нимодипин увеличивает возбудимость пирамидных нейронов CA1 кролика в зависимости от возраста и концентрации. J. Neurophysiol. 68 , 2100–2109 (1992).
CAS PubMed Google ученый
Тибо, О. и Ландфилд, П. В. Увеличение одиночных кальциевых каналов L-типа в нейронах гиппокампа во время старения. Наука 272 , 1017–1020 (1996). Первое свидетельство того, что увеличение активированного напряжением притока Ca 2+ в старых нейронах гиппокампа CA1 связано с возрастным увеличением Ca 2+ каналов L-типа.
CAS PubMed Google ученый
Тоеску, Э.С., Верхратский, А. и Ландфилд, П.В. Ca 2+ регуляция и экспрессия генов при нормальном старении мозга. Trends Neurosci. 27 , 614–620 (2004).
CAS PubMed Google ученый
Фостер Т. и Норрис К. М. Возрастные изменения в процессах, зависимых от Ca 2+ : связь с синаптической пластичностью гиппокампа. Гиппокамп 7 , 602–612 (1997).
CAS PubMed Google ученый
Landfield, P. W. Нейробиологические механизмы гиппокампа возрастной дисфункции памяти. Neurobiol. Старение 9 , 571–579 (1988).
CAS PubMed Google ученый
Томбо, Г. К., Роу, В. Б. и Роуз, Г. М. Медленная постгиперполяризация нейронов CA1 гиппокампа зависит от способности к пространственному обучению у старых крыс Fisher 344. J. Neurosci. 25 , 2609–2616 (2005).
CAS PubMed Google ученый
Барнс, К. А., Макнотон, Б. Л. и О’Киф, Дж. Потеря специфичности места в спайк-клетках гиппокампа стареющих крыс. Neurobiol. Старение 4 , 113–119 (1983).
CAS PubMed Google ученый
Маркус, Э.Дж., Барнс, К. А., Макнотон, Б. Л., Гладден, В. Л. и Скэггс, В. Е. Пространственное информационное наполнение и надежность нейронов CA1 гиппокампа: эффекты визуального ввода. Гиппокамп 4 , 410–421 (1994).
CAS PubMed Google ученый
Мизумори, С. Дж., Лавуа, А. М. и Каляни, А. Перераспределение пространственного представления в гиппокампе старых крыс, выполняющих задачу пространственной памяти. Behav. Neurosci. 110 , 1006–1016 (1996).
CAS PubMed Google ученый
Барнс, К. А., Састер, М. С., Шен, Дж. И Макнотон, Б. Л. Мультистабильность когнитивных карт в гиппокампе старых крыс. Nature 388 , 272–275 (1997). Первое сообщение об изменении стабильности карт места гиппокампа у старых крыс, которое коррелирует с бимодальной эффективностью старых крыс при выполнении пространственной версии задания Морриса по плаванию.
CAS PubMed Google ученый
Шен, Дж., Барнс, К. А., Макнотон, Б. Л., Скэггс, В. Э. и Уивер, К. Л. Влияние старения на зависимую от опыта пластичность клеток места гиппокампа. J. Neurosci. 17 , 6769–6782 (1997). Многие клетки места гиппокампа регистрировали одновременно у молодых и старых крыс, чтобы выявить, что механизмы пластичности, вызванной поведением, у старых крыс дефектны.
CAS PubMed Google ученый
Танила, Х., Шапиро, М., Галлахер, М. и Эйхенбаум, Х. Старение мозга: изменения в природе кодирования информации гиппокампом. J. Neurosci. 17 , 5155–5166 (1997).
CAS PubMed Google ученый
Смит, А. К., Джеррард, Дж. Л., Барнс, К. А. и Макнотон, Б.L. Влияние возраста на взрывные характеристики пирамидных клеток гиппокампа крысы. Нейроотчет 11 , 3865–3871 (2000).
CAS PubMed Google ученый
Олер Дж. А. и Маркус Э. Дж. Возрастной дефицит способности кодировать изменение контекста: анализ ячеек места. Гиппокамп 10 , 338–350 (2000).
CAS PubMed Google ученый
Уилсон, И.A., Ikonen, S., Gallagher, M., Eichenbaum, H. & Tanila, H. Возрастные изменения в клетках места являются субрегиональными. J. Neurosci. 25 , 6877–6886 (2005). Первый документ, в котором описывается различие между влиянием старения на сети CA1 и CA3. Это проясняет, что эти области не могут быть объединены при сравнении возраста.
CAS PubMed Google ученый
Чанг, Ю.M., Rosene, D.L., Killiany, R.J., Mangiamele, L.A. & Luebke, J. I. Повышенная скорость возбуждения потенциала действия пирамидных клеток слоя 2/3 в префронтальной коре в значительной степени связана с когнитивными функциями у старых обезьян. Cereb. Cortex 15 , 409–418 (2005).
PubMed Google ученый
Бондарефф В. и Гейнисман Ю. Потеря синапсов в зубчатой извилине стареющей крысы. Am.J. Anat. 145 , 129–136 (1976).
CAS PubMed Google ученый
Гейнисман Ю., Бондарефф В. и Додж Дж. Т. Частичная деафферентация нейронов в зубчатой извилине стареющей крысы. Brain Res. 134 , 541–545 (1977).
CAS PubMed Google ученый
Гейнисман, Ю., де Толедо-Моррелл, Л.И Моррелл, Ф. Потеря перфорированных синапсов в зубчатой извилине: морфологический субстрат дефицита памяти у старых крыс. Proc. Natl Acad. Sci. USA 83 , 3027–3031 (1986).
CAS PubMed Google ученый
Гейнисман, Ю., де Толедо-Моррелл, Л., Моррелл, Ф., Персина, И.С. и Росси, М. Возрастная потеря аксоспинных синапсов, образованных двумя афферентными системами в зубчатой извилине крысы, как показано методом объективного стереологического диссектора. Гиппокамп 2 , 437–444 (1992).
CAS PubMed Google ученый
Барнс, К. А., Рао, Г. и Хьюстон, Ф. П. Изменение порога индукции LTP у старых крыс в синапсе перфорантного пути и гранулярных клеток. Neurobiol. Старение 21 , 613–620 (2000).
CAS PubMed Google ученый
Фостер, Т. К., Барнс, К. А., Рао, Г. и Макнотон, Б. Л. Увеличение квантового размера перфорантного пути у старых крыс F-344. Neurobiol. Старение 12 , 441–448 (1991).
CAS PubMed Google ученый
Барнс, К. А., Рао, Г. и Макнотон, Б. Л. Повышенное электротоническое взаимодействие в гиппокампе старых крыс: возможный механизм изменений клеточной возбудимости. J. Comp. Neurol. 259 , 549–558 (1987).
CAS PubMed Google ученый
Гейнисман, Ю. и др. Старение, пространственное обучение и общее количество синапсов в stratum radiatum СА1 крысы. Neurobiol. Старение 25 , 407–416 (2004).
CAS PubMed Google ученый
Николсон, Д. А., Йошида, Р., Берри, Р. В., Галлахер, М. и Гейнисман, Ю. Уменьшение размера перфорированных постсинаптических плотностей в аксоспинозных синапсах гиппокампа и возрастные нарушения пространственного обучения. J. Neurosci. 24 , 7648–7653 (2004).
CAS PubMed Google ученый
Landfield, P. W., Pitler, T. A. и Applegate, M. D. Влияние высоких отношений Mg 2+ к Ca 2+ на частотную потенциацию в срезах гиппокампа молодых и старых крыс. J. Neurophysiol. 56 , 797–811 (1986).
CAS PubMed Google ученый
Деупре, Д.Л., Брэдли, Дж. И Тернер, Д. А. Возрастные изменения потенциации в области CA1 у крыс F344. Neurobiol. Старение 14 , 249–258 (1993).
CAS PubMed Google ученый
Barnes, C. A., Rao, G. & Shen, J. Возрастное снижение опосредованного N -метил-D-аспартата R возбуждающего постсинаптического потенциала в области CA1 гиппокампа. Neurobiol. Старение 18 , 445–452 (1997).
CAS PubMed Google ученый
Розенцвейг, Э. С., Рао, Г., Макнотон, Б. Л. и Барнс, К. А. Роль временного суммирования в возрастных долгосрочных дефицитах индукции потенцирования. Гиппокамп 7 , 549–558 (1997).
CAS PubMed Google ученый
Барнс, К. А., Рао, Г. и Орр, Г. Связанное с возрастом снижение ВПСП, вызванного Шаффером, у бодрствующих, свободно ведущих крыс. Neural Plast. 7 , 167–178 (2000).
CAS PubMed PubMed Central Google ученый
Томбо, Г. К., Роу, В. Б., Чоу, А. Р., Майкл, Т. Х. и Роуз, Г. М. Тета-частотное синаптическое потенцирование в CA1 in vitro. отличает когнитивно нарушенных крыс от старых крыс Fischer 344 без нарушений. J. Neurosci. 22 , 9932–9940 (2002).
CAS PubMed Google ученый
Блисс, Т.В. и Коллингридж, Г. Л. Синаптическая модель памяти: долговременная потенциация в гиппокампе. Nature 361 , 31–39 (1993).
CAS PubMed PubMed Central Google ученый
Малинов, Р. и Маленка, Р. С. Транспортировка рецепторов AMPA и синаптическая пластичность. Annu. Rev. Neurosci. 25 , 103–126 (2002).
CAS PubMed Google ученый
Диана, Г., Доменичи, М. Р., Лоиццо, А., Скотти де Каролис, А. и Сагрателла, С. Различия в возрасте и деформации в обучении крыс в месте обучения и потенциации частоты зубчатых извилин гиппокампа. Neurosci. Lett. 171 , 113–116 (1994).
CAS PubMed Google ученый
Диана, Г., Скотти де Каролис, А., Франк, К., Доменичи, М. Р. и Сагрателла, С. Избирательное снижение потенцирования зубной частоты гиппокампа у старых крыс с нарушением обучения местам. Brain Res. Бык. 35 , 107–111 (1994).
CAS PubMed Google ученый
Landfield, P. W. & Lynch, G. Нарушение моносинаптической потенциации в in vitro срезах гиппокампа старых крыс с недостаточной памятью. J. Gerontol. 32 , 523–533 (1977).
CAS PubMed Google ученый
Лэндфилд, П.W., McGaugh, J. L. и Lynch, G. Нарушение процессов синаптической потенциации в гиппокампе у старых крыс с недостаточной памятью. Brain Res. 150 , 85–101 (1978).
CAS PubMed Google ученый
Dieguez, D. Jr & Barea-Rodriguez, E. J. Старение нарушает позднюю фазу долговременной потенциации в медиальном перфорантном пути — синапсе CA3 у бодрствующих крыс. Synapse 52 , 53–61 (2004).
CAS PubMed PubMed Central Google ученый
Барнс, К. А., Рао, Г. и Хьюстон, Ф. П. Изменение порога индукции LTP у старых крыс в синапсе перфорантного пути и гранулярных клеток. Neurobiol. Старение 21 , 613–620 (2000).
CAS PubMed Google ученый
Барнс, К. А., Рао, Г. и Макнотон, Б. Л. Функциональная целостность NMDA-зависимых механизмов индукции LTP на протяжении всей жизни крыс F-344. Учиться. Mem. 3 , 124–137 (1996).
CAS PubMed Google ученый
Deupree, D. L., Turner, D. A. и Watters, C. L. Пространственная характеристика коррелирует с потенциацией in vitro у молодых и старых крыс Fischer 344. Brain Res. 554 , 1–9 (1991).
CAS PubMed Google ученый
Мур, К.I., Browning, M. D. & Rose, G. M. Пластичность гиппокампа, индуцированная примированным взрывом, но не долговременной потенциацией, стимуляцией нарушается в области CA1 у старых крыс Fischer 344. Гиппокамп 3 , 57–66 (1993). В противоположность более ранним сообщениям об индукции интактного гиппокампа LTP (см. Ссылки 38 и 83), результаты этого исследования предполагают, что задействование механизмов индукции пластичности около порогового значения становится более трудным с возрастом.
CAS PubMed Google ученый
Медведь, м.Ф., Купер, Л. Н. и Эбнер, Ф. Ф. Физиологические основы теории модификации синапсов. Наука 237 , 42–48 (1987).
CAS PubMed Google ученый
Медведь М. Ф. и Маленка Р. С. Синаптическая пластичность: LTP и LTD. Curr. Мнение. Neurobiol. 4 , 389–399 (1994).
CAS PubMed Google ученый
Тибо, О., Hadley, R. & Landfield, P. W. Повышенная постсинаптическая активность [Ca 2+ ] i и кальциевых каналов L-типа в старых нейронах гиппокампа: связь с нарушенной синаптической пластичностью. J. Neurosci. 21 , 9744–9756 (2001).
CAS PubMed Google ученый
Норрис, К.М., Корол, Д.Л. и Фостер, Т.С. Повышенная восприимчивость к индукции долговременной депрессии и долговременная реверсия потенцирования во время старения. J. Neurosci. 16 , 5382–5392 (1996). Представляет первую характеристику гомосинаптического обращения LTD / LTP у старых крыс и показывает, что пластичность, вызванная низкочастотной стимуляцией, увеличивается во время старения, вероятно, в результате нарушения регуляции Ca 2+ .
CAS PubMed Google ученый
Kumar, A. & Foster, T. C. Внутриклеточные запасы кальция способствуют повышенной чувствительности к индукции LTD во время старения. Brain Res. 1031 , 125–128 (2005).
CAS PubMed Google ученый
Морган, Дж. И., Коэн, Д. Р., Хемпстед, Дж. Л. и Карран, Т. Картирование паттернов экспрессии c-fos в центральной нервной системе после приступа. Наука 237 , 192–197 (1987).
CAS Google ученый
Коул, А. Дж., Саффен, Д.У., Барабан, Дж. М. и Уорли, П. Ф. Быстрое увеличение информационной РНК немедленного раннего гена в нейронах гиппокампа за счет активации синаптического рецептора NMDA. Nature 340 , 474–476 (1989).
CAS PubMed Google ученый
Dragunow, M. et al. Долгосрочная потенциация и индукция мРНК и белков c-fos в зубчатой извилине неанестезированных крыс. Neurosci. Lett. 101 , 274–280 (1989).
CAS PubMed Google ученый
Wisden, W. et al. Дифференциальная экспрессия немедленных ранних генов в гиппокампе и спинном мозге. Нейрон 4 , 603–614 (1990).
CAS PubMed Google ученый
Гузовски, Дж. Ф., Макнотон, Б. Л., Барнс, К. А. и Уорли, П. Ф. Экспрессия немедленного-раннего гена Arc , специфичная для окружающей среды, в нейрональных ансамблях гиппокампа. Nature Neurosci. 2 , 1120–1124 (1999).
CAS PubMed Google ученый
Платеник, Дж., Курамото, Н. и Йонеда, Ю. Молекулярные механизмы, связанные с длительной консолидацией сигналов NMDA. Life Sci. 67 , 335–364 (2000).
CAS PubMed Google ученый
Клейтон, Д. Ф.Геномный потенциал действия. Neurobiol. Учиться. Mem. 74 , 185–216 (2000).
CAS PubMed Google ученый
Jones, M. W. et al. Необходимость в непосредственном раннем гене Zif268 в экспрессии поздних ДП и долгосрочных воспоминаний. Nature Neurosci. 4 , 289–296 (2001).
CAS Google ученый
Рети, И.М., Редди, Р., Уорли, П. Ф. и Барабан, Дж. М. Заметная экспрессия Narp в проекционных путях и конечных полях. J. Neurochem. 82 , 935–944 (2002).
CAS PubMed Google ученый
О’Брайен, Р. Дж. И др. Синаптическая кластеризация рецепторов AMPA внеклеточным продуктом предраннего гена Narp. Нейрон 23 , 309–323 (1999).
CAS PubMed Google ученый
Стюард, О., Wallace, C. S., Lyford, G. L. & Worley, P. F. Синаптическая активация заставляет мРНК для IEG Arc селективно локализоваться вблизи активированных постсинаптических сайтов на дендритах. Нейрон 21 , 741–751 (1998).
CAS PubMed Google ученый
Lyford, G. L. et al. Arc, фактор роста и ген, регулируемый активностью, кодирует новый белок, связанный с цитоскелетом, который обогащен нейрональными дендритами. Нейрон 14 , 433–445 (1995).
CAS PubMed Google ученый
Guzowski, J. F. et al. Ингибирование зависимой от активности экспрессии белка дуги в гиппокампе крыс ухудшает поддержание долгосрочной потенциации и консолидацию долговременной памяти. J. Neurosci. 20 , 3993–4001 (2000).
CAS PubMed Google ученый
Цзян, К.Х., Цзянь, Дж. З., Шульц, П. Г. и Ху, Ю. Влияние старения на экспрессию генов в гипоталамусе и коре головного мозга мышей. Proc. Natl Acad. Sci. США 98 , 1930–1934 (2001).
CAS PubMed Google ученый
Ли, К. К., Вайндрух, Р. и Пролла, Т. А. Профиль экспрессии генов стареющего мозга мышей. Nature Genet. 25 , 294–297 (2000).
CAS PubMed Google ученый
Блэлок, Э.M. et al. Генные микромассивы при старении гиппокампа: статистическое профилирование выявляет новые процессы, коррелированные с когнитивными нарушениями. J. Neurosci. 23 , 3807–3819 (2003). Отчет о первом эксперименте с микрочипами экспрессии генов на крысах, характеризуемых поведением, который показывает возрастное изменение уровней экспрессии нескольких генов в состоянии покоя, которое коррелирует со снижением когнитивных функций.
CAS PubMed Google ученый
Смит Д.Р., Хойт, Э. С., Галлахер, М., Швабе, Р. Ф. и Лунд, П. К. Влияние возраста и когнитивного статуса на базальный уровень активности AP-1 в гиппокампе крыс. Neurobiol. Старение 22 , 773–786 (2001).
CAS PubMed Google ученый
Ланахан, А., Лайфорд, Г., Стивенсон, Г. С., Уорли, П. Ф. и Барнс, С. А. Избирательное изменение транскрипционного ответа, вызванного длительной потенциацией, в гиппокампе старых крыс с нарушенной памятью. J. Neurosci. 17 , 2876–2885 (1997).
CAS PubMed Google ученый
Смолл, С. А., Чавла, М. К., Буонокор, М., Рапп, П. Р. и Барнс, К. А. Визуализация коррелятов функции мозга у обезьян и крыс позволяет выделить субрегион гиппокампа, дифференциально уязвимый к старению. Proc. Natl Acad. Sci. США 101 , 7181–7186 (2004). Авторы использовали разные методы визуализации у крыс и обезьян и сообщают о межвидовом консенсусе, что зубчатая извилина гиппокампа особенно уязвима для воздействия старения.Эти результаты согласуются с предыдущим сообщением о людях.
CAS PubMed Google ученый
Смолл, С. А., Цай, В. Ю., ДеЛапаз, Р., Мейе, Р. и Стерн, Ю. Визуализация функции гиппокампа на протяжении всей жизни человека: снижение памяти является нормальным или нет? Ann. Neurol. 51 , 290–295 (2002).
PubMed Google ученый
Кентрос, К.и другие. Устранение долговременной стабильности новых карт мест гиппокампа путем блокады рецепторов NMDA. Наука 280 , 2121–2126 (1998).
CAS PubMed Google ученый
Экстром, А. Д., Мельцер, Дж., МакНотон, Б. Л. и Барнс, К. А. Антагонизм к рецепторам NMDA блокирует зависимое от опыта расширение «полей мест» гиппокампа. Нейрон 31 , 631–638 (2001).
CAS PubMed Google ученый
О’Киф, Дж.& Достровский Дж. Гиппокамп как пространственная карта. Предварительные свидетельства активности единиц у свободно передвигающейся крысы. Brain Res. 34 , 171–175 (1971).
CAS PubMed Google ученый
Юнг, М. В. и Макнотон, Б. Л. Пространственная избирательность активности единиц в гранулярном слое гиппокампа. Гиппокамп 3 , 165–182 (1993).
CAS PubMed PubMed Central Google ученый
Уилсон, М.А. и Макнотон, Б. Л. Динамика кода гиппокампа для пространства. Наука 261 , 1055–1058 (1993).
CAS PubMed Google ученый
МакНотон, Б. Л., Барнс, К. А. и О’Киф, Дж. Вклад положения, направления и скорости в активность отдельных единиц в гиппокампе свободно движущихся крыс. Exp. Brain Res. 52 , 41–49 (1983).
CAS PubMed Google ученый
Мюллер Р.У., Босток, Э., Таубе, Дж. С. и Куби, Дж. Л. О свойствах направленного возбуждения пространственных клеток гиппокампа. J. Neurosci. 14 , 7235–7251 (1994).
CAS PubMed Google ученый
Markus, E.J. et al. Взаимодействие между местоположением и задачей влияет на пространственное и направленное возбуждение нейронов гиппокампа. J. Neurosci. 15 , 7079–7094 (1995).
CAS PubMed Google ученый
Мехта, М.Р., Барнс, К. А. и Макнотон, Б. Л. Зависимое от опыта асимметричное расширение полей мест гиппокампа. Proc. Natl Acad. Sci. USA 94 , 8918–8921 (1997).
CAS PubMed Google ученый
Хебб Д. Организация поведения: нейрофизиологическая теория (Уайли, Нью-Йорк, 1949).
Google ученый
Томпсон, Л.Т. и Бест, П. Дж. Долговременная стабильность пространственно-полевой активности отдельных единиц, записанная в спинном гиппокампе свободно ведущих крыс. Brain Res. 509 , 299–308 (1990).
CAS PubMed Google ученый
Агнихотри, Н. Т., Хокинс, Р. Д., Кандел, Э. Р. и Кентрос, К. Долговременная стабильность новых полей мест гиппокампа требует синтеза нового белка. Proc. Natl Acad.Sci. США 101 , 3656–3661 (2004).
CAS PubMed Google ученый
Танила, Х., Сипила, П., Шапиро, М. и Эйхенбаум, Х. Старение мозга: нарушение кодирования новых сигналов окружающей среды. J. Neurosci. 17 , 5167–5174 (1997).
CAS PubMed Google ученый
Ли, И., Йоганарасимха, Д., Рао, Г.& Knierim, J. J. Сравнение популяционной когерентности клеток места в субполях гиппокампа CA1 и CA3. Природа 430 , 456–459 (2004).
CAS PubMed Google ученый
Leutgeb, S., Leutgeb, J. K., Treves, A., Moser, M. B. & Moser, E. I. Отчетливые ансамблевые коды в областях CA3 и CA1 гиппокампа. Наука 305 , 1295–1298 (2004).
CAS PubMed Google ученый
Ваздарьянова, А.И Гузовски, Дж. Ф. Различия в ответах популяции нейронов гиппокампа на модификации окружающего контекста: доказательства различных, но взаимодополняющих функций ансамблей CA3 и CA1. J. Neurosci. 24 , 6489–6496 (2004).
CAS PubMed Google ученый
Марр Д. Простая память: теория архикортекса. Фил. Пер. R. Soc. Лондон. В 262 , 23–81 (1971).
CAS Google ученый
МакНотон, Б. Л. и Моррис, Р. Г. Улучшение синапсов в гиппокампе и хранение информации в системе распределенной памяти. Trends Neurosci. 10 , 408–415 (1987).
Google ученый
Спенсер, В. Д. и Раз, Н. Дифференциальные эффекты старения на память для содержания и контекста: метаанализ. Psychol. Старение 10 , 527–539 (1995).
CAS PubMed Google ученый
Макинтайр, Дж. С. и Крейк, Ф. И. Возрастные различия в памяти для информации об элементе и источнике. Банка. J. Psychol. 41 , 175–192 (1987).
CAS PubMed Google ученый
Вилкнисс, С. М., Джонс, М. Г., Король, Д. Л., Голд, П. Э. и Мэннинг, К. А. Возрастные различия в экологически обоснованном исследовании изучения маршрута. Psychol. Старение 12 , 372–375 (1997).
CAS PubMed Google ученый
Ньюман М. и Кашнаик А. Пространственная память и старение: производительность на человеческом аналоге водного лабиринта Морриса. Aging Neuropsychol. Cogn. 7 , 86–93 (2000). Подобно крысам и обезьянам, в сухой версии задания Морриса по плаванию у здоровых пожилых людей нарушено запоминание местоположения ориентира по отношению к сигналам помещения.
Google ученый
Лай, З. К., Мосс, М. Б., Киллиани, Р. Дж., Розен, Д. Л. и Херндон, Дж. Дж. Дисфункция исполнительной системы у пожилых обезьян: пространственное обучение и обучение с обращением объектов. Neurobiol. Старение 16 , 947–954 (1995).
CAS PubMed Google ученый
Рапп П. Р., Кански М. Т. и Робертс Дж. А. Нарушение обработки пространственной информации у старых обезьян с сохраненной памятью распознавания. Нейроотчет 8 , 1923–1928 (1997). Эти результаты показывают существенное соответствие между данными о крысах и обезьянах, показывая, что у пожилых животных нарушены задачи, проверяющие пространственную память.
CAS PubMed Google ученый
Head, E. et al. Пространственное обучение и память как функция возраста у собаки. Behav. Neurosci. 109 , 851–858 (1995).
CAS PubMed Google ученый
Марковская, А.L. et al. Индивидуальные различия в старении: поведенческие и нейробиологические корреляты. Neurobiol. Старение 10 , 31–43 (1989).
CAS PubMed Google ученый
Галлахер М. и Рапп П. Р. Использование моделей на животных для изучения влияния старения на познание. Annu. Rev. Psychol. 48 , 339–370 (1997).
CAS PubMed Google ученый
Бах, М.E. et al. Возрастные дефекты пространственной памяти коррелируют с дефектами поздней фазы долгосрочной потенциации гиппокампа in vitro и ослабляются лекарствами, которые усиливают сигнальный путь цАМФ. Proc. Natl Acad. Sci. США 96 , 5280–5285 (1999).
CAS PubMed Google ученый
Розенцвейг, Э. С. и Барнс, К. А. Влияние старения на функцию гиппокампа: пластичность, сетевая динамика и познание. Прог. Neurobiol. 69 , 2003, 143–179.
CAS PubMed Google ученый
Кристиан, К. М. и Томпсон, Р. Ф. Нейронные субстраты кондиционирования моргания: получение и удержание. Учиться. Mem. 10 , 427–455 (2003).
PubMed Google ученый
Кишимото Ю., Судзуки М., Кавахара С.& Кирино, Ю. Возрастозависимое нарушение задержки и отслеживания моргания у мышей. Нейроотчет 12 , 3349–3352 (2001).
CAS PubMed Google ученый
Кнуттинен, М. Г., Гамелли, А. Э., Вайс, К., Пауэр, Дж. М. и Дистерхофт, Дж. Ф. Возрастные эффекты на кондиционирование моргания у гибридных крыс F344 × BN F1. Neurobiol. Старение 22 , 1–8 (2001).
CAS PubMed Google ученый
Томпсон, Л.Т., Мойер, Дж. Р. Младший и Дистерхофт, Дж. Ф. Тренировка моргания глаз у кроликов демонстрирует неоднородность обучаемости как между возрастными группами, так и внутри них. Neurobiol. Старение 17 , 619–629 (1996).
CAS PubMed Google ученый
Соломон, П. Р. и Грокчиа-Эллисон, М. Э. Классическое кондиционирование у старых кроликов: кондиционирование с задержкой, отслеживанием и с длительной задержкой. Behav. Neurosci. 110 , 427–435 (1996).
CAS PubMed Google ученый
Финкбинер, Р. Г. и Вудрафф-Пак, Д. С. Классическое кондиционирование моргания в зрелом возрасте: влияние возраста и интервала между стимулами на приобретение в парадигме следа. Psychol. Старение 6 , 109–117 (1991).
CAS PubMed Google ученый
Фунахаши, С., Брюс, К. Дж. И Гольдман-Ракич, П.S. Дорсолатеральные префронтальные поражения и задержка реакции глазодвигательного аппарата: доказательства мнемонических «скотом». J. Neurosci. 13 , 1479–1497 (1993).
CAS PubMed Google ученый
Мэйр, Р. Г., Берк, Дж. А. и Портер, М. С. Поражения лобной коры, гиппокампа и интраламинарных ядер таламуса по-разному влияют на запоминание у крыс. Behav. Neurosci. 112 , 772–792 (1998).
CAS PubMed Google ученый
Годфруа О., Кабаре М., Пети-Шеналь В., Пруво Дж. П. и Руссо М. Управляющие функции лобных долей. Модульность центрально-надзорной системы? Cortex 35 , 1–20 (1999).
CAS PubMed Google ученый
Даннет, С. Б., Эвенден, Дж. Л. и Иверсен, С.D. Зависящий от задержки дефицит кратковременной памяти у старых крыс. Психофармакология (Berl.) 96 , 174–180 (1988).
CAS Google ученый
Мосс, М. Б., Розен, Д. Л. и Петерс, А. Влияние старения на зрительную память распознавания у макак-резусов. Neurobiol. Старение 9 , 495–502 (1988).
CAS PubMed Google ученый
Рапп, П.Р. и Амарал, Д. Г. Доказательства дисфункции памяти в зависимости от задачи у пожилых обезьян. J. Neurosci. 9 , 3568–3576 (1989).
CAS PubMed Google ученый
Мосс, М. Б., Киллиани, Р. Дж., Лай, З. К., Розен, Д. Л. и Херндон, Дж. Г. Интервал памяти распознавания у макак-резусов пожилого возраста. Neurobiol. Старение 18 , 13–19 (1997).
CAS PubMed Google ученый
Лайонс-Уоррен, А., Lillie, R. & Hershey, T. Краткосрочные и долгосрочные характеристики пространственной задержки отклика на протяжении всей жизни. Dev. Neuropsychol. 26 , 661–678 (2004).
PubMed Google ученый
Wiig, K. A. & Burwell, R. D. Ухудшение памяти при выполнении задачи отсроченного несоответствия с положением после повреждения периринальной коры у крысы. Behav. Neurosci. 112 , 827–838 (1998).
CAS PubMed Google ученый
Buffalo, E.A. et al. Диссоциация между эффектами повреждения периринальной коры и области TE. Учиться. Mem. 6 , 572–599 (1999).
CAS PubMed PubMed Central Google ученый
Rhodes, M. G. Возрастные различия в результатах теста сортировки карточек в Висконсине: метааналитический обзор. Psychol. Старение 19 , 482–494 (2004). Если рассматривать образовательный статус и методику тестирования, по сравнению с более молодыми людьми, пожилые люди демонстрируют дефицит показателей исполнительной функции, оцениваемой с помощью задачи сортировки карт Висконсина, что коррелирует с возрастным снижением функционирования префронтальной коры.
PubMed Google ученый
Мур, Т. Л., Киллиани, Р. Дж., Херндон, Дж. Г., Розен, Д. Л. и Мосс, М. Б. Нарушение абстракции и смена установки у старых макак-резусов. Neurobiol. Старение 24 , 125–134 (2003). По сравнению с молодыми взрослыми обезьянами, пожилые обезьяны страдают нарушением функции животного аналога задачи сортировки карт Висконсина, что предполагает возрастное снижение функционирования префронтальной коры.Это согласуется с исследованиями на людях.
PubMed Google ученый
Паксинос, Г. и Уотсон, К. Мозг крысы в стереотаксических координатах 4-е изд. (Academic, San Diego, 1998).
Google ученый
Gothard, K. M., Skaggs, W. E., Moore, K. M. & McNaughton, B.L. Привязка нейронной активности CA1 гиппокампа к нескольким опорным кадрам в навигационной задаче на основе ориентиров. J. Neurosci. 16 , 823–835 (1996).
CAS PubMed Google ученый
Макнотон, Б. Л., О’Киф, Дж. И Барнс, К. А. Стереотрод: новый метод одновременного выделения нескольких отдельных единиц в центральной нервной системе из нескольких записей. J. Neurosci. Методы 8 , 391–397 (1983).
CAS PubMed Google ученый
Серый, C.М., Мальдонадо, П. Э., Уилсон, М. и МакНотон, Б. Тетроды заметно повышают надежность и эффективность многокомпонентной изоляции от многокомпонентных записей в полосатом теле кошки. J. Neurosci. Методы 63 , 43–54 (1995).
CAS PubMed Google ученый
Рао, Г., Барнс, К. А. и Макнотон, Б. Л. Внутриклеточное флуоресцентное окрашивание карбоксифлуоресцеином: быстрый и надежный метод количественного определения связывания красителя в центральной нервной системе млекопитающих. J. Neurosci. Методы 16 , 251–263 (1986).
CAS PubMed Google ученый